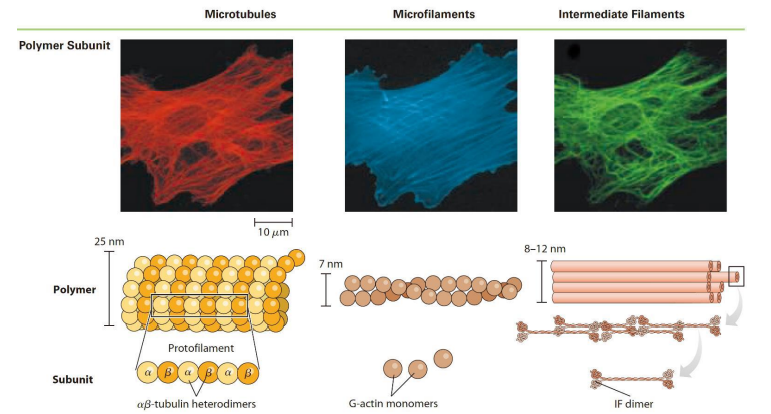
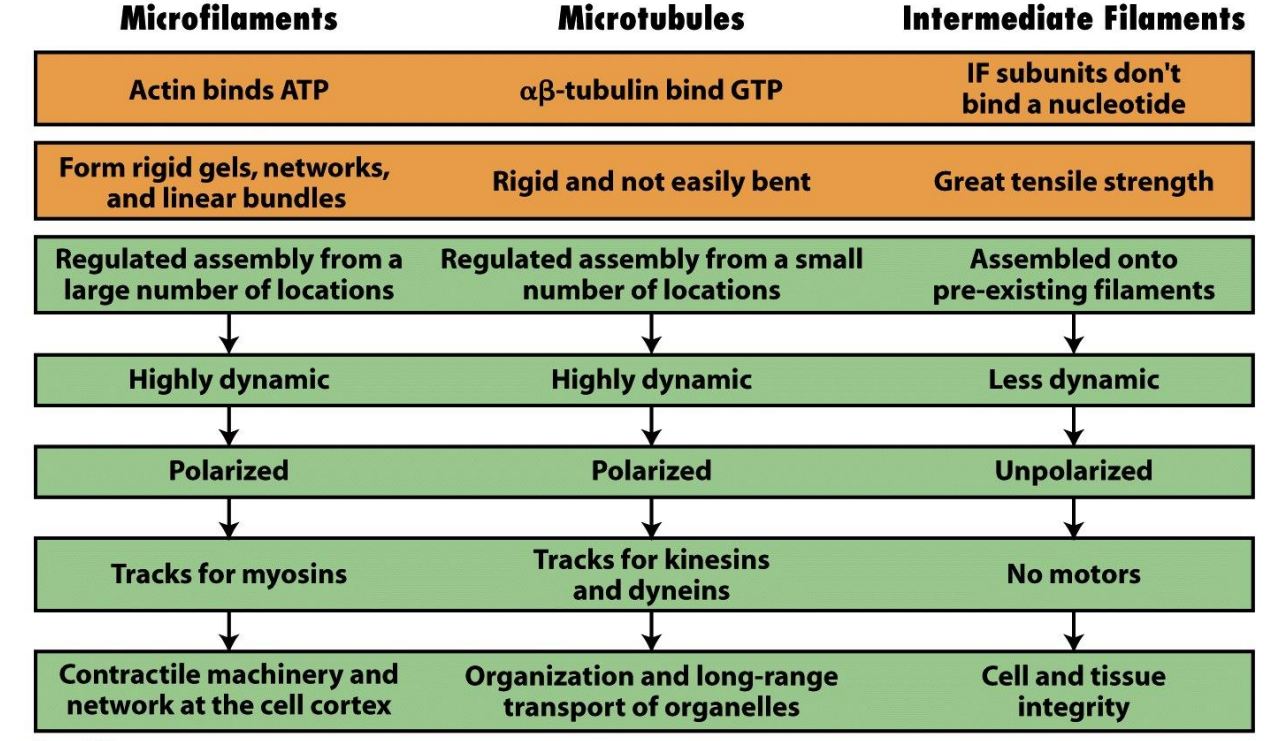
Eukaryotes Have Three Basic Types of Cytoskeletal Elements(微管、微丝、中间纤维)
Bacteria Have Cytoskeletal Systems 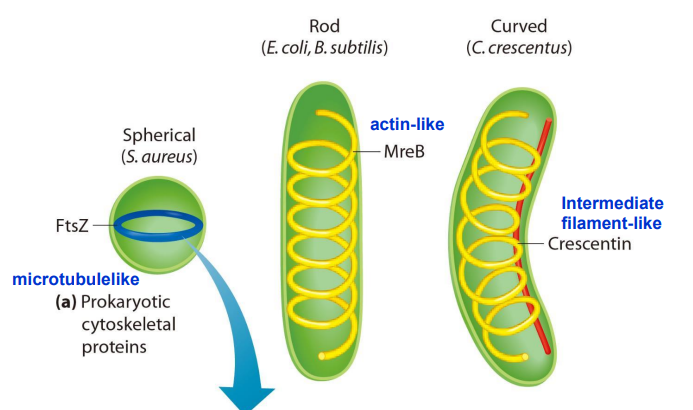
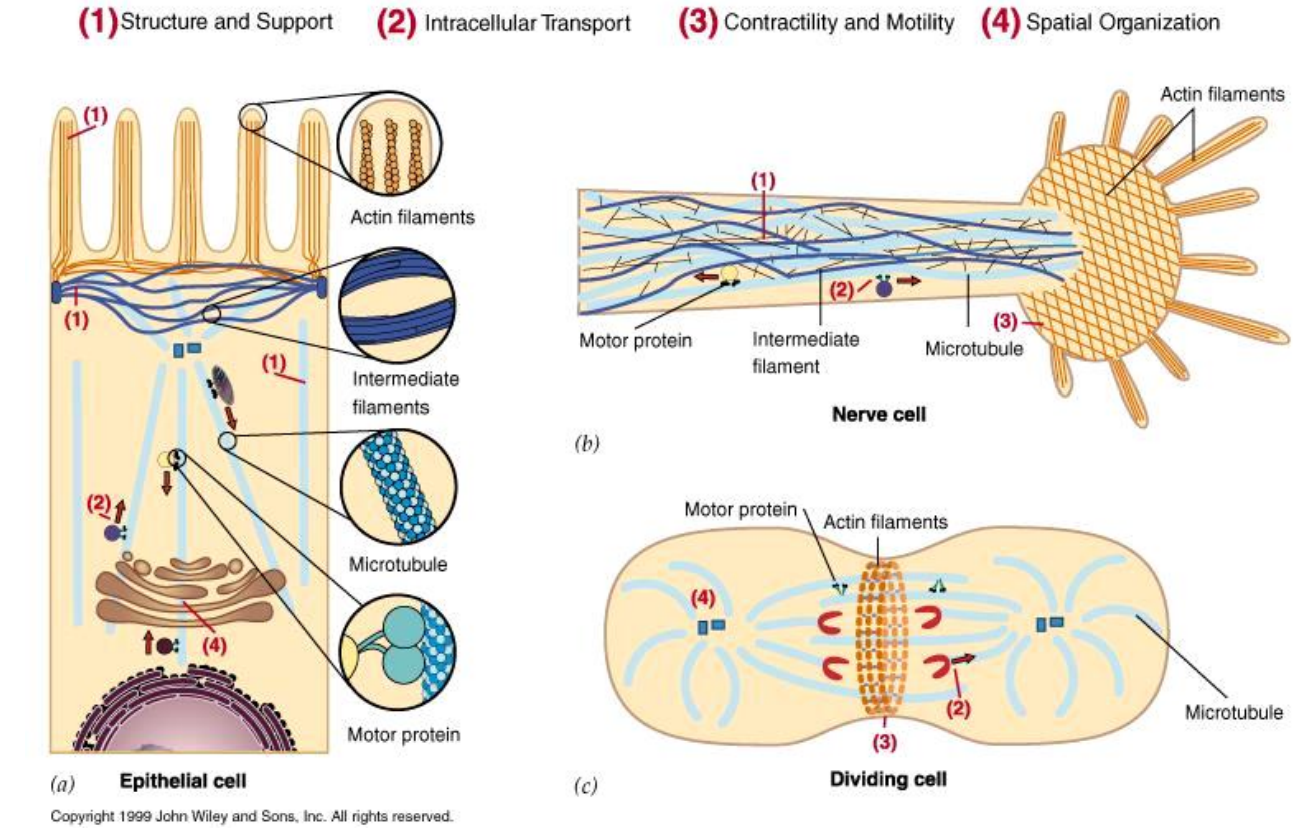
Key to Cytoskeletal Functions
目录
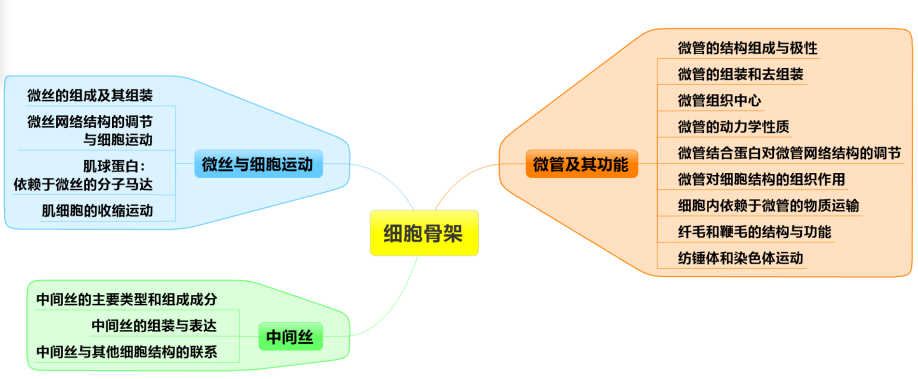
0.细胞骨架的基本概念
第一节 微丝与细胞运动
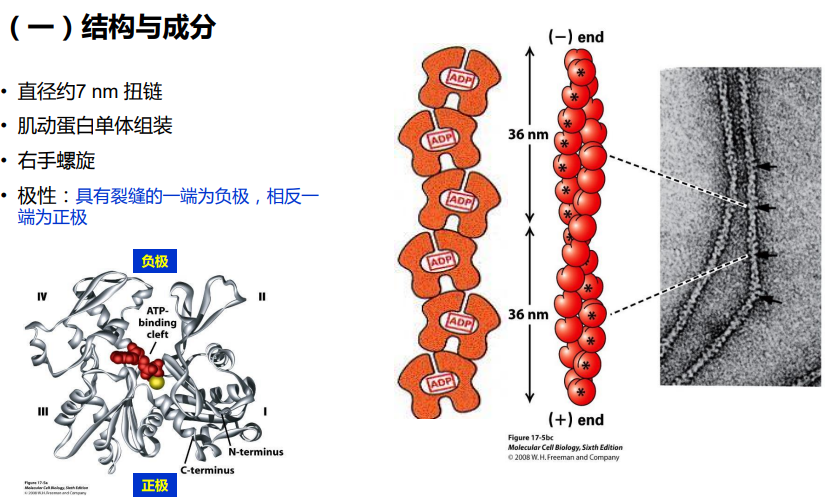
微丝(microfilament, MF)
ATP帽(ATP Cap)与踏车行为(treadmilling)
微丝细胞内组装:受微丝结合蛋白影响
(三)影响微丝组装的特异性药物
二、微丝网络结构的调节与细胞运动
(一)非肌肉细胞内微丝的结合蛋白
(二)细胞皮层(cell cortex)
(三)应力纤维
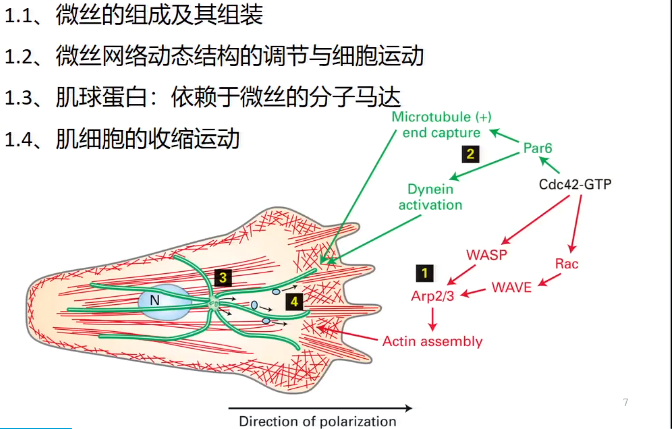
(四)细胞伪足的形成与细胞迁移
三、肌球蛋白:依赖于微丝的分子马达
(一)肌球蛋白的种类
(二)肌球蛋白的结构
1. Ⅱ型肌球蛋白:传统的肌球蛋白(conventional myosin)
2. 其他类型的肌球蛋白(unconventional myosin)
四、肌细胞的收缩运动
(一)肌纤维的结构
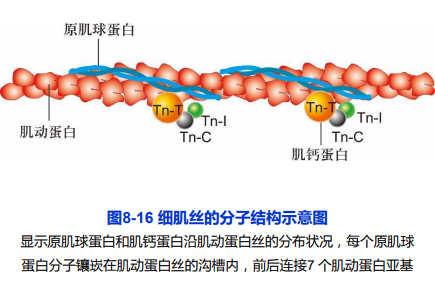
细肌丝结构
肌肉收缩系统中的其他蛋白组分
(二)肌肉收缩的滑动模型
滑行学说(sliding theory)
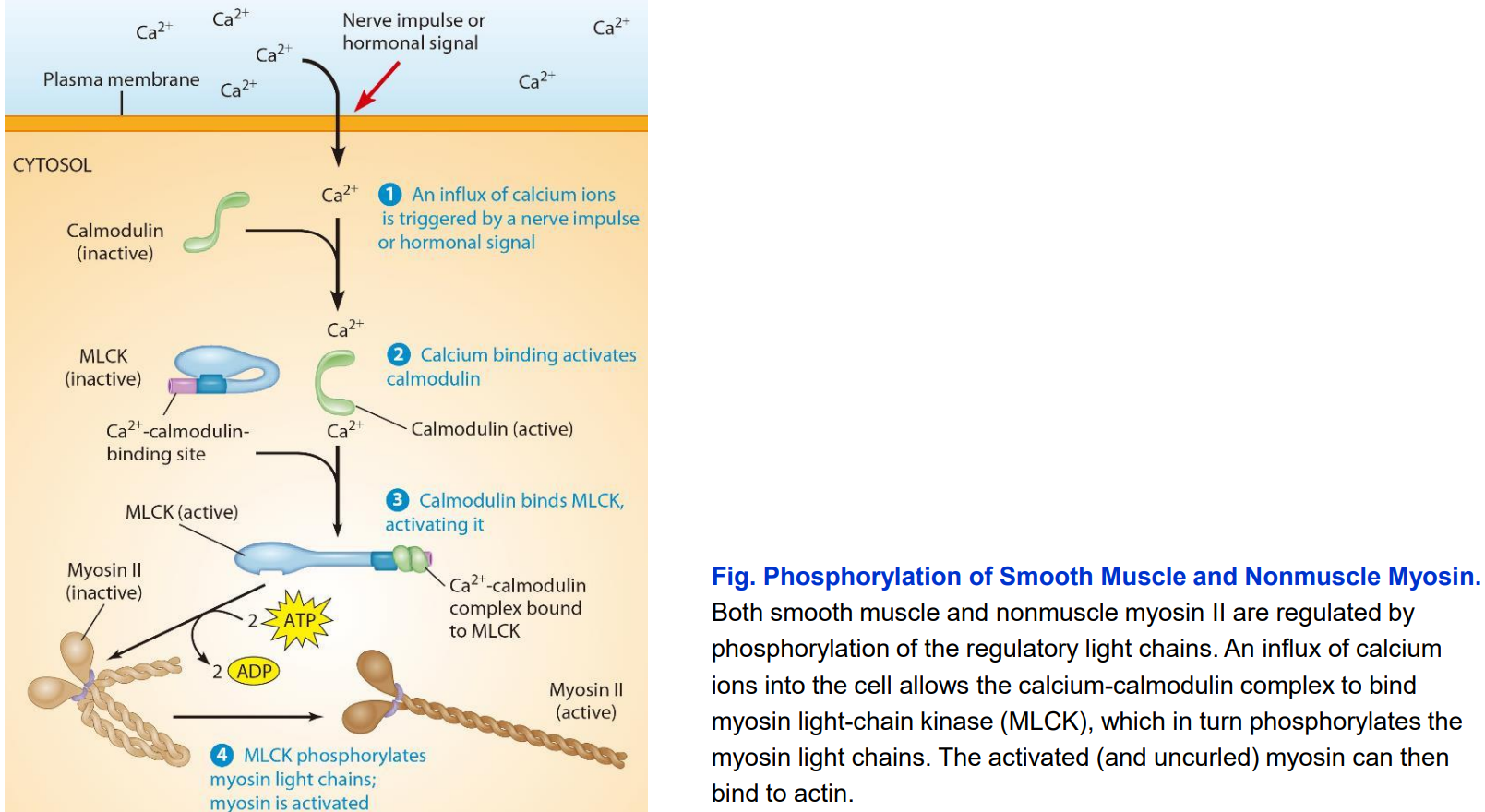
Phosphorylation of myosin II by myosin light-chain kinase (MLCK)
第二节 微管及功能
一、微管的结构组成与极性
细胞内微管的3种类型
二、微管的组装和去组装
(一)微管的体外组装与踏车行为
(二)作用于微管的特异性药物
三、微管组织中心(MTOC)体内组装
(一)中心体
(二)基体和其他微管组织中心
四、微管的动力学性质
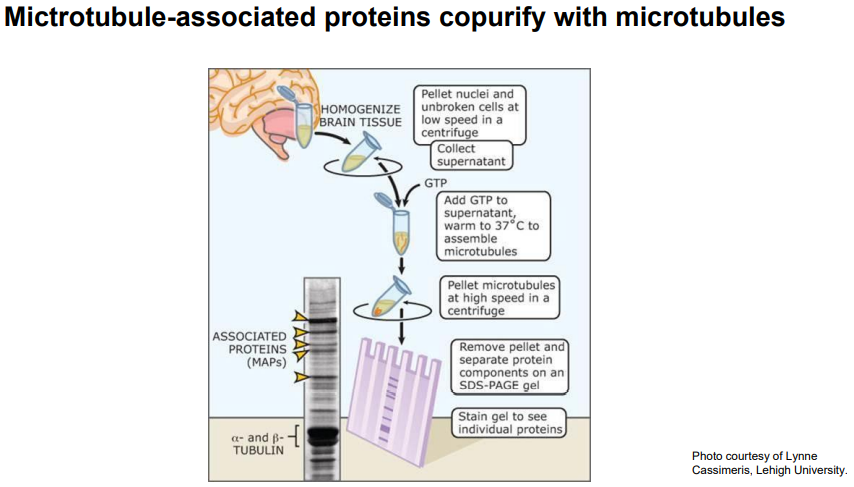
五、微管结合蛋白对微管网络结构的调节
六、微管对细胞结构的组织作用
七、细胞内依赖于微管的物质运输
(一)驱动蛋白
驱动蛋白家族成员的结构与功能
2. 驱动蛋白沿微管运动的分子机制
“步行” 模型和“尺蠖” 爬行模型
“步 行”(hand over hand) 模型
(二)胞质动力蛋白及其功能
轴丝动力蛋白 axonemal dynein
Movement of pigment granules: color adjustment
八、纤毛和鞭毛的结构与功能
(一)纤毛的结构及组装
(二)纤毛或鞭毛的运动机制
细菌鞭毛运动机制
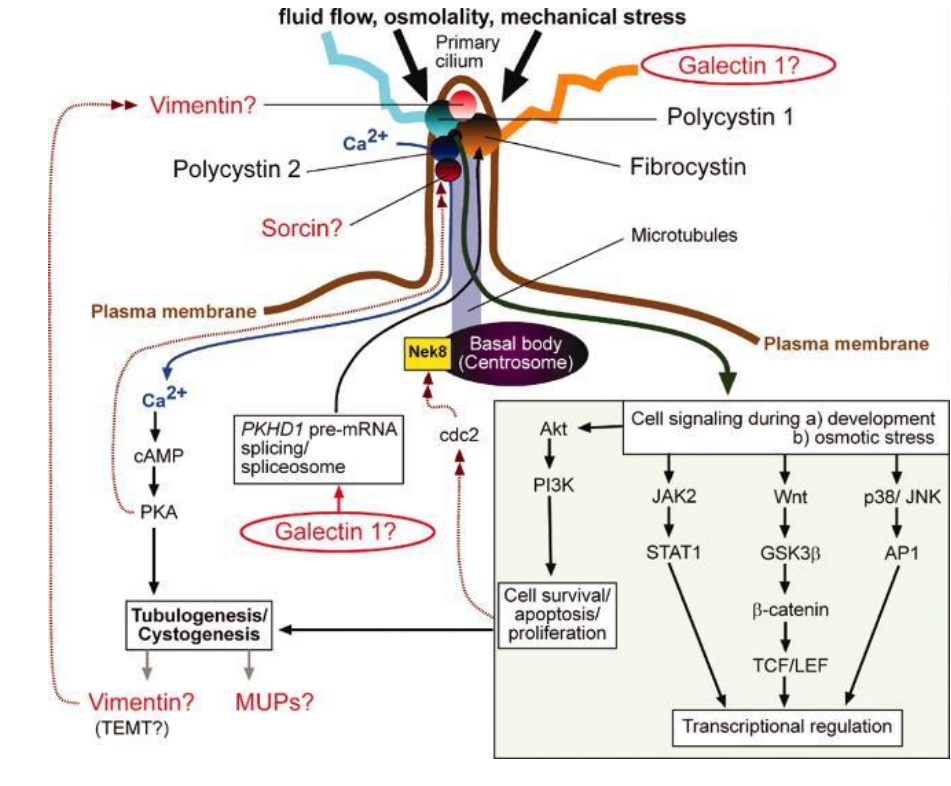
(三)纤毛的功能
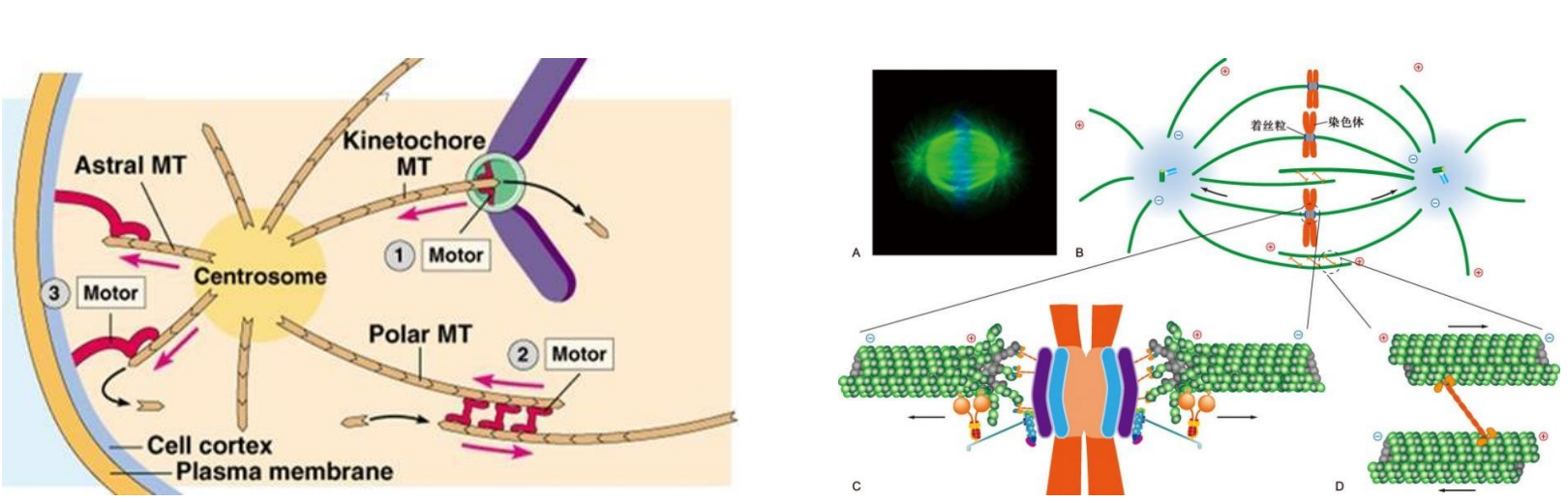
九、纺锤体和染色体运动
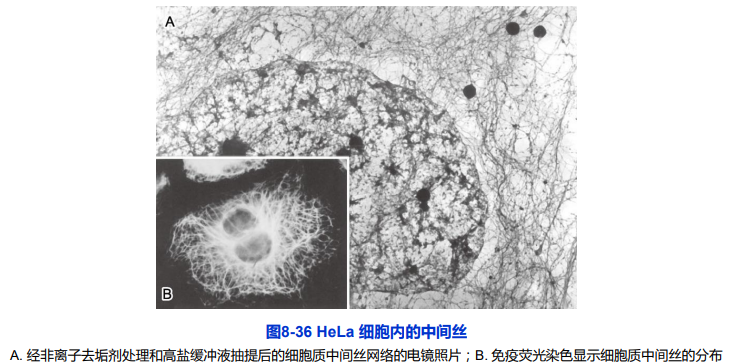
第三节 中间丝
中间丝又称中间纤维(intermediate filament,IF)
一、中间丝的主要类型和组成成分
中间丝蛋白分子结构
二、中间丝的组装与表达
三、中间丝与其他细胞结构的联系
中间丝结合蛋白
中间丝的功能
单纯性疱性表皮松懈症(simplex epidermolysis bullosa)
本章内容提要
0.细胞骨架的基本概念
细胞骨架(cytoskeleton)一般指真核细胞细胞质内由蛋白质组成的复杂纤维状网架结构体系,包括:微丝、微管、中间纤维。
广义的细胞骨架还包括核骨架和膜骨架。
细胞骨架是高度动态的结构体系,对细胞的结构和功能发挥组织作用,并进一步影响细胞的形态、运动、胞内物质运输及周围的细胞和环境。
第一节 微丝与细胞运动
微丝(microfilament, MF)
微丝又称作肌动蛋白丝(actin filament)或纤维状肌动蛋白(fibrous actin ,F-actin)
直径7 nm;存在于所有的真核细胞中;组装/去组装与多种细胞生命活动密切相关。
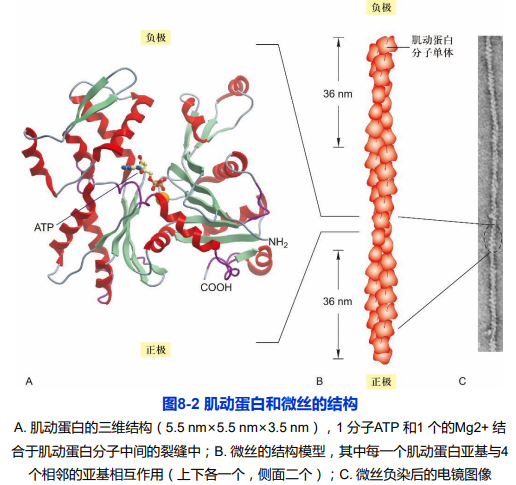
肌动蛋白(actin)由一条多肽链构成的球形蛋白质,是组成微丝的基本蛋白质,分子量为43kDa,序列
Eukaryotes Have Three Basic Types of Cytoskeletal Elements(微管、微丝、中间纤维)
Bacteria Have Cytoskeletal Systems
Key to Cytoskeletal Functions
0.细胞骨架的基本概念
细胞骨架(cytoskeleton)一般指真核细胞细胞质内由蛋白质组成的复杂纤维状网架结构体系,包括:微丝、微管、中间纤维。
广义的细胞骨架还包括核骨架和膜骨架。
细胞骨架是高度动态的结构体系,对细胞的结构和功能发挥组织作用,并进一步影响细胞的形态、运动、胞内物质运输及周围的细胞和环境。
第一节 微丝与细胞运动
微丝(microfilament, MF)
微丝又称作肌动蛋白丝(actin filament)或纤维状肌动蛋白(fibrous actin ,F-actin)
直径7 nm;存在于所有的真核细胞中;组装/去组装与多种细胞生命活动密切相关。
肌动蛋白(actin)由一条多肽链构成的球形蛋白质,是组成微丝的基本蛋白质,分子量为43kDa,序列高度保守,具有组织和表达阶段的特异性。存在ATP/ADP结合位点;
α- 肌动蛋白为横纹肌、心肌、血管平滑肌和肠道平滑肌所特有;
β- 肌动蛋白和 γ-肌动蛋白存在于所有的细胞中
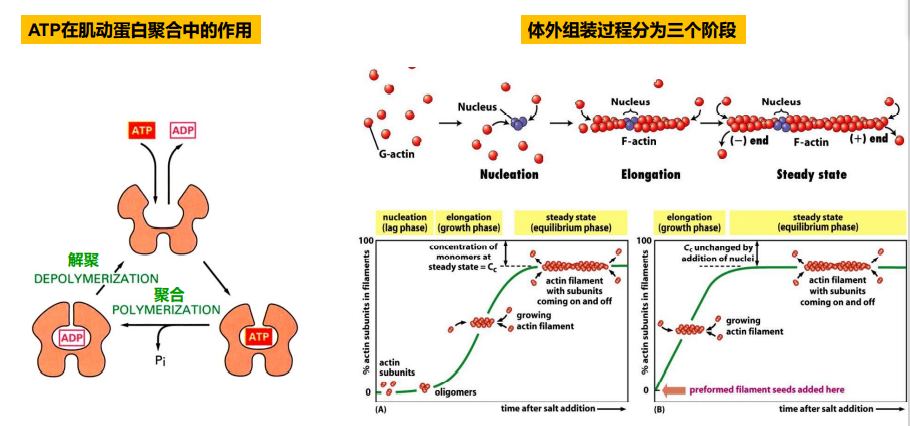
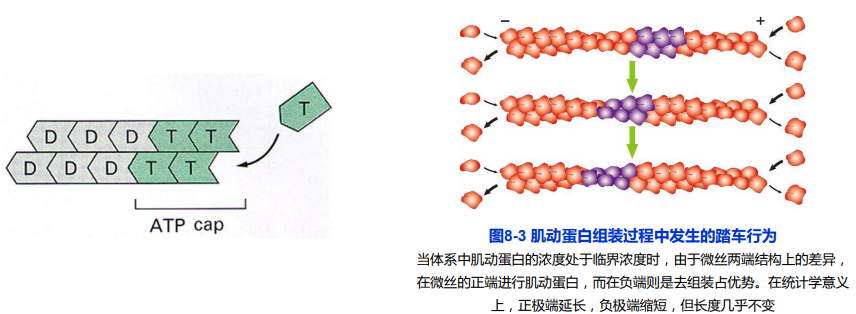
ATP帽(ATP Cap)与踏车行为(treadmilling)
当微丝末端聚合速度大于ATP水解速度时,会在末端形成一个ATP帽;
踏车行为:在体外组装过程中,微丝正极由于肌动蛋白亚基不断增长而延长,负极由于肌动蛋白亚基去组装而缩短的现象。
微丝细胞内组装:受微丝结合蛋白影响
(三)影响微丝组装的特异性药物
细胞松弛素(cytochalasin)
- 与微丝结合之后将微丝切断,并结合在微丝末端阻碍肌动蛋白的聚合,但对微丝解聚没有影响;
- 破坏微丝网络结构,并阻止细胞的运动。
鬼笔环肽(phalloidin)
- 与微丝表面有强亲和力,不与肌动蛋白单体结合;
- 阻止微丝的解聚,使其保持稳定状态。
二、微丝网络结构的调节与细胞运动
细胞中大多数微丝结构处于动态的组装和去组装过程,并通过这种方式实现功能。
(一)非肌肉细胞内微丝的结合蛋白
1.肌动蛋白单体结合蛋白
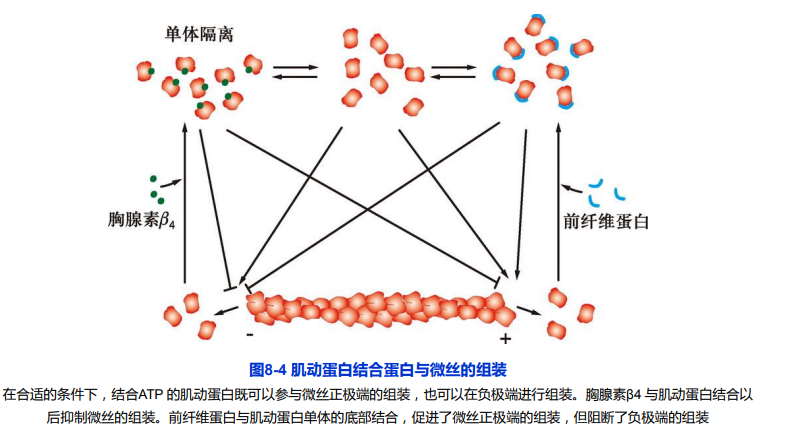
结合ATP的肌动蛋白既可以参与微丝正极端的组装,也可以在负极端进行组装。胸腺素β4与肌动蛋白结合以后抑制微丝的组装。前纤维蛋白与肌动蛋白单体的底部结合,促进了微丝正极端的组装,但阻断了负极端的组装。
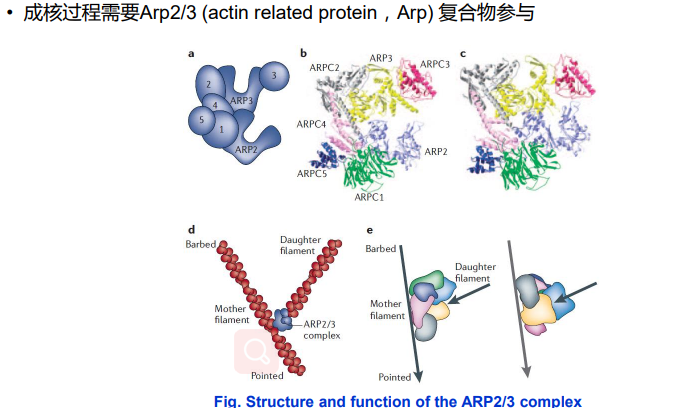
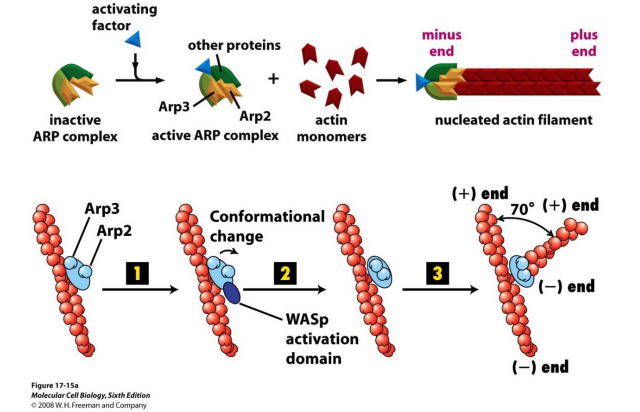
2. 成核蛋白-Arp2/3促进微丝蛋白的分叉
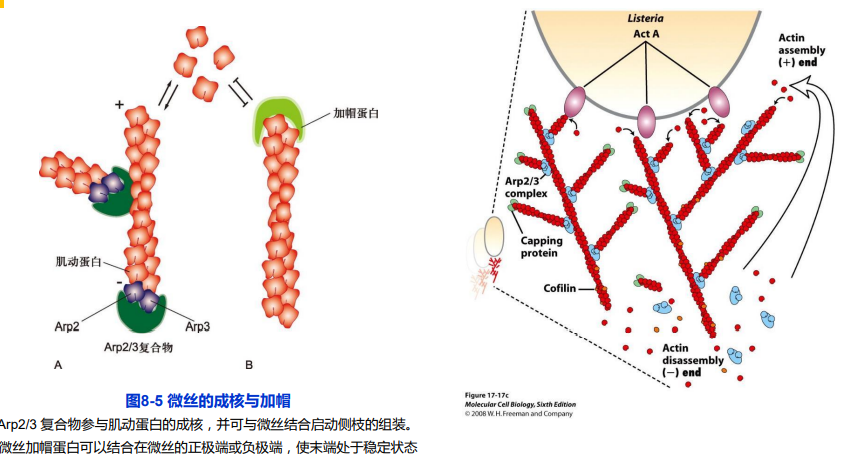
3. 加帽蛋白(capping protein)-增加稳定性
4. 交联蛋白
- 成束蛋白(bunding protein):将相邻的微丝交联成平行排列状态;
- 凝胶形成蛋白(gel-forming protein):将微丝联接成网状。
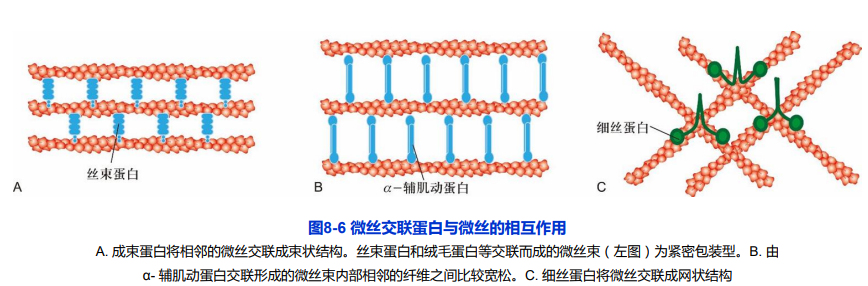
5. 割断及解聚蛋白
- 凝溶胶蛋白(gelsolin):在高Ca2+(>1 μmol/L)情况下 能将较长微丝切断,使肌动蛋白由凝胶态转化成溶胶态
- 丝切蛋白/ 肌动蛋白解聚因子(cofilin/actin depolymerizing factor,Cofilin/ADF):能与游离的肌动 蛋白或微丝结合,提高微丝的解聚速度
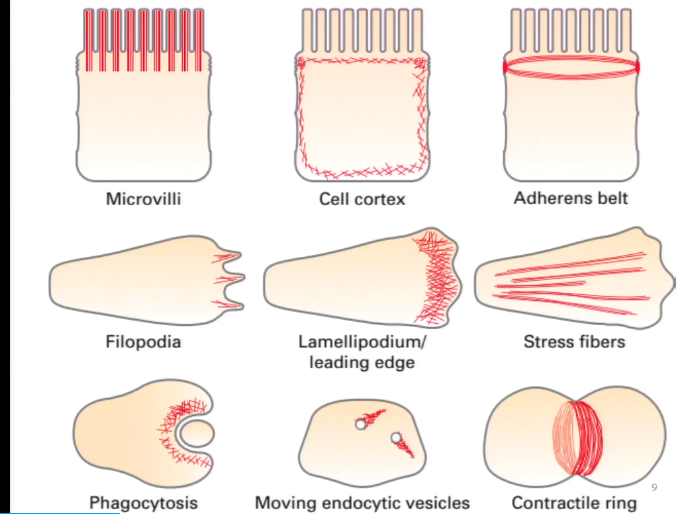
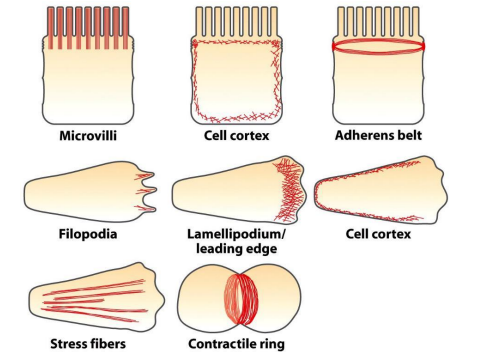
(二)细胞皮层(cell cortex)
细胞内大部分微丝都集中在紧贴细胞质膜的细胞质区域,并由微丝交联蛋白交联成凝胶态的三维网络结构。
限制膜蛋白的流动性;为细胞质膜提供强度和韧性;维持细胞形状;细胞的多种运动,如胞质环流(cyclosis)、阿米巴运动(amoiboid)、变皱膜运动 (ruffled membrane locomotion)、 吞噬(phagocytosis)以及膜蛋白的定位等与皮层内肌动蛋白的溶胶态- 凝胶态转化相关。
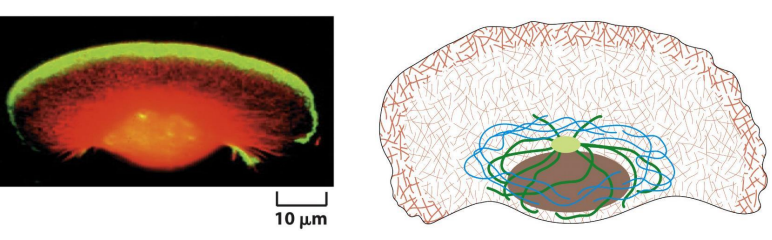
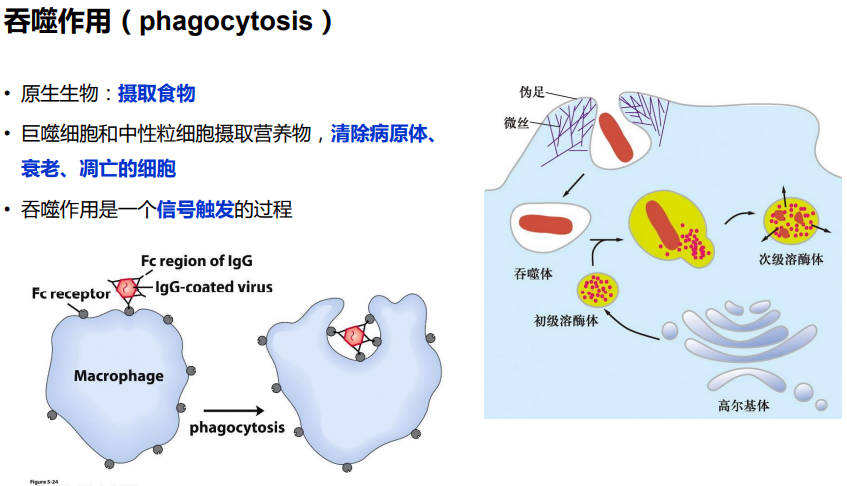
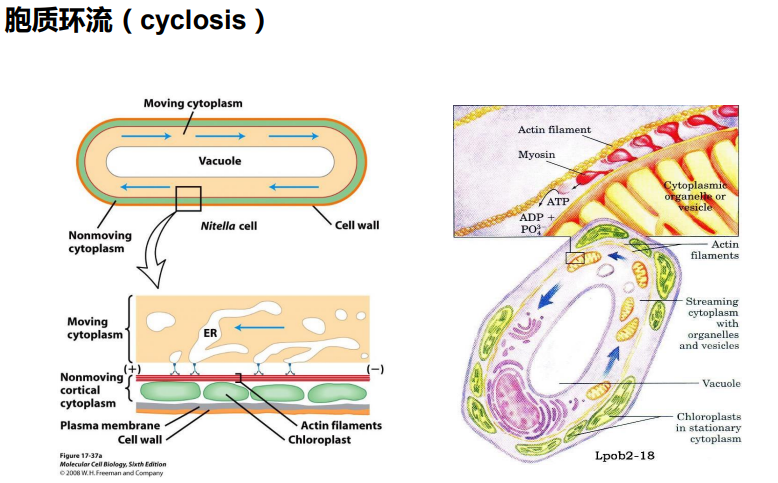
(三)应力纤维
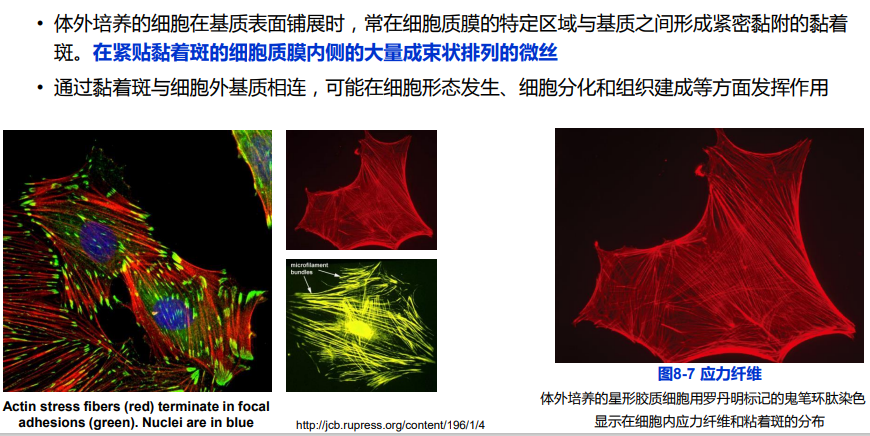
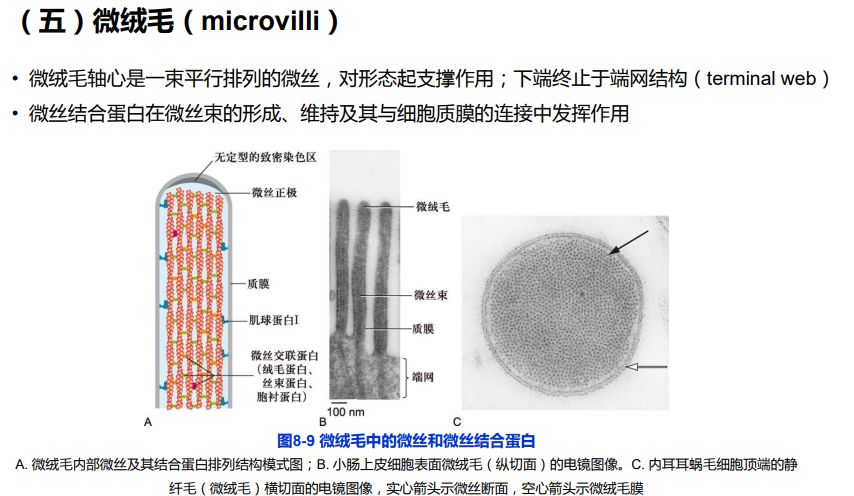
(四)细胞伪足的形成与细胞迁移

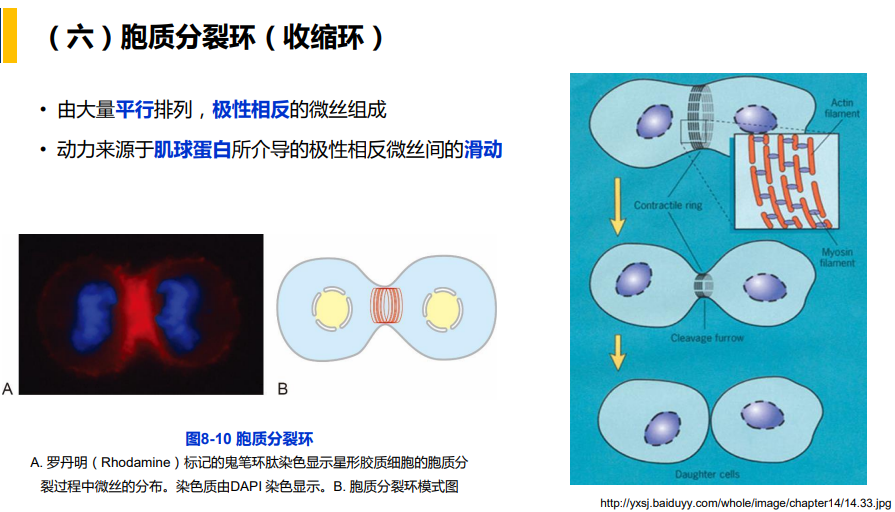
三、肌球蛋白:依赖于微丝的分子马达
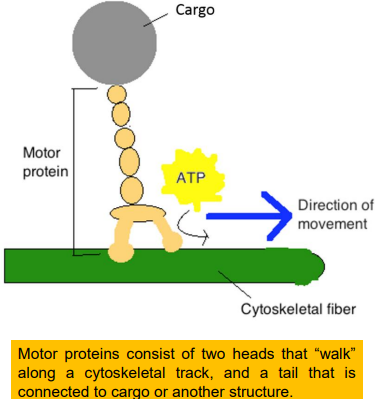
马达蛋白可分为三种:
- 沿微丝运动的肌球蛋白(myosin);
- 沿微管移动的驱动蛋白(kinesin);
- 沿微管移动的动力蛋白(dynein);
马达结构域与微丝或微管结合;货物结合结构域与膜性细胞器或大分子复合物特异性结合;
能量转换:利用水解ATP所提供的能量沿微丝或微管运动。
(一)肌球蛋白的种类
- 骨骼肌细胞内,多个Ⅱ型肌球蛋白分子组装成肌原纤维的粗肌丝;
- 马达结构域是肌球蛋白超家族成员最保守的部位,是肌球蛋白定性和分类的依据;
- 基于马达结构域多肽链一级结构的相似性,至少可以将肌球蛋白超家族成员分成18种家族;
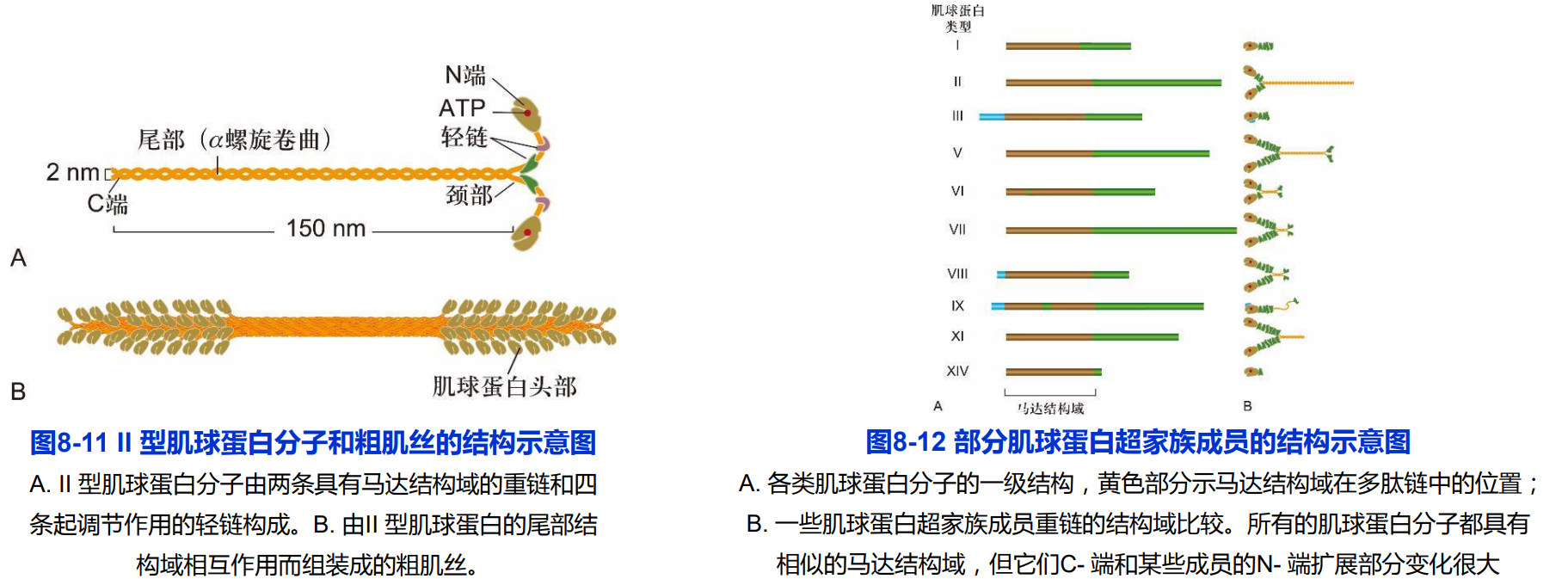
(二)肌球蛋白的结构
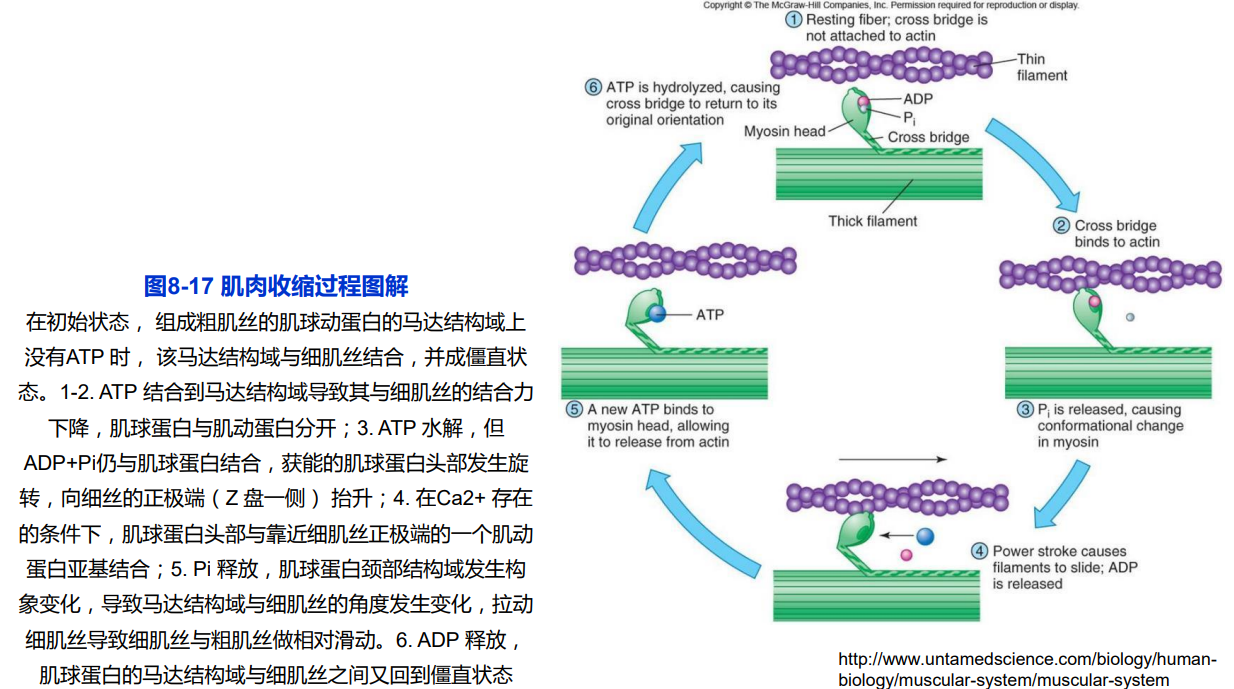
肌球蛋白是沿着微丝运动的马达蛋白,通常有三个功能结构域;
- 马达结构域:位于肌球蛋白的头部,包含一个肌动蛋白亚基结合位点和一个具有ATP酶活性的ATP结合位点,负责将ATP水解释放发化学能转变为机械能。
- 调控结构域:连接马达结构域和尾部杆状区的一段α螺旋,轻链的结合部位,发挥杠杆作用。
- 尾部结构域:选择性与所运输的“货物”结合;
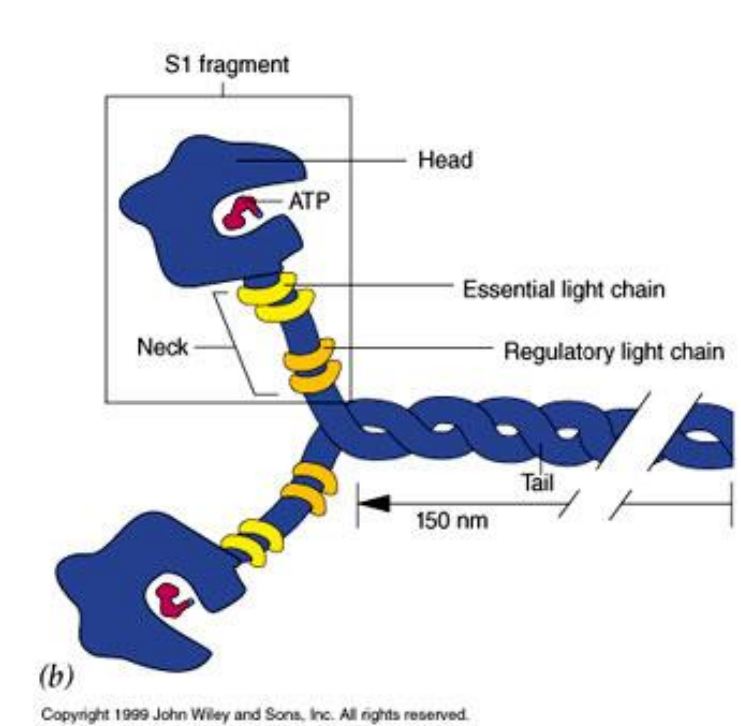
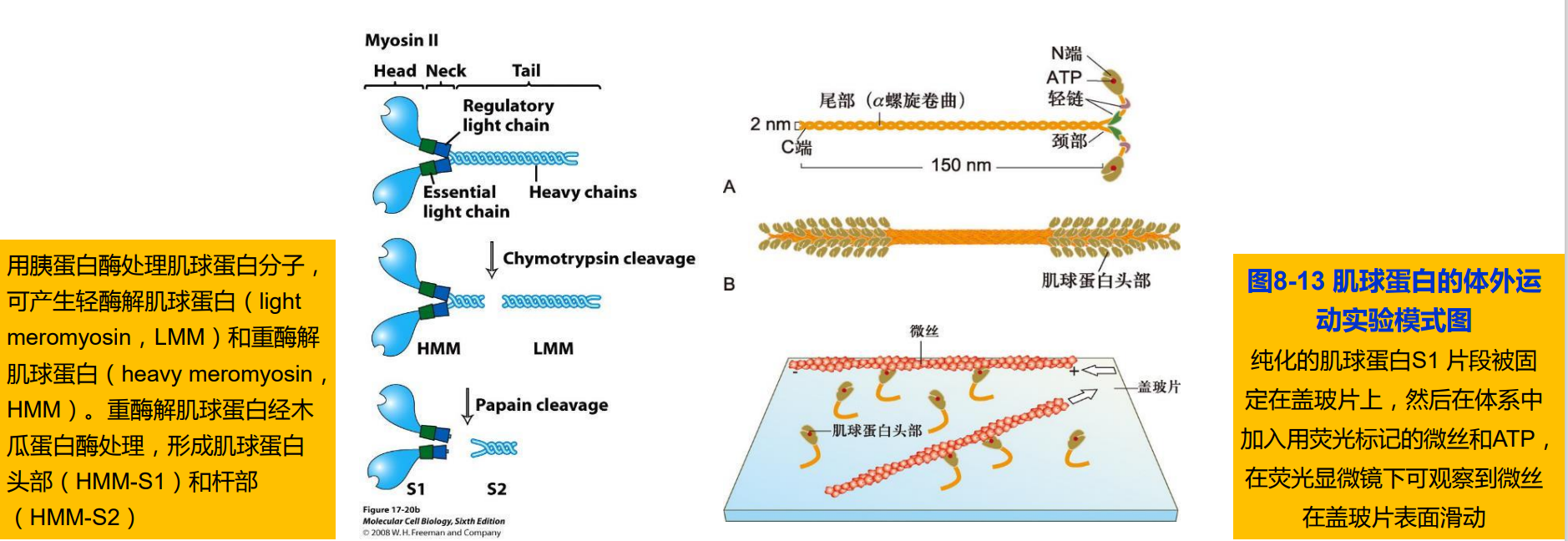
1. Ⅱ型肌球蛋白:传统的肌球蛋白(conventional myosin)
- 肌细胞:组装成肌原纤维的粗丝;
- 非肌细胞:胞质分裂过程中收缩环的主要结构成分;应力纤维的结构成分;
- 包含两条重链和4条轻链,形成一个高度不对称的结构。
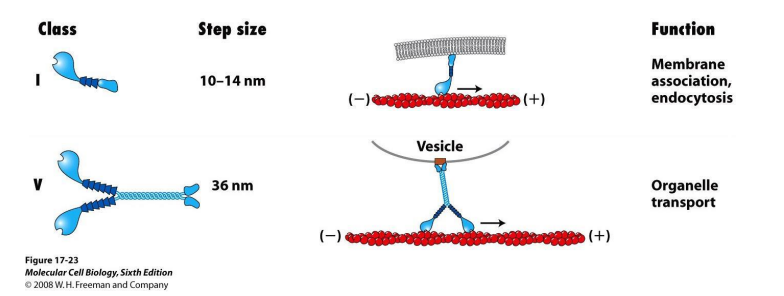
2. 其他类型的肌球蛋白(unconventional myosin)
- Ⅰ型肌球蛋白分子:一个头部(马达结构域)和一个尾部
- Ⅴ型肌球蛋白分子:两条肽链组成的二聚体;两个头部交替与微丝结合
四、肌细胞的收缩运动
(一)肌纤维的结构
- 肌纤维:肌原纤维(myofibrils);
- 肌节(sarcomere);
- 粗肌丝;细肌丝;
- 横桥;
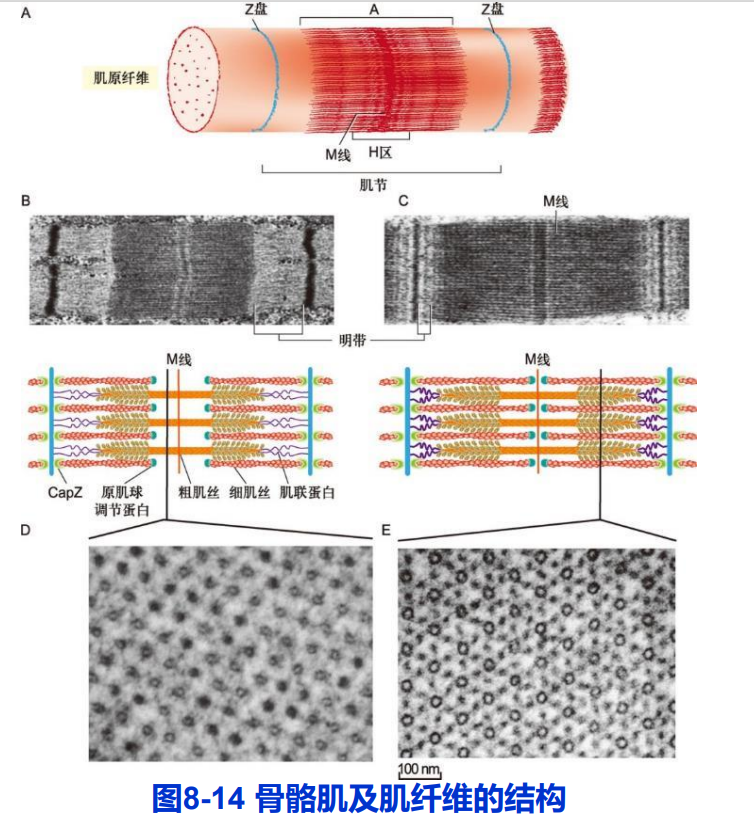
细肌丝结构
肌动蛋白;
- 原肌球蛋白(tropomyosin,Tm);
- 两条平行多肽形成α螺旋构型,调节actin与myosin头部结合;
肌钙蛋白(troponin,Tn):
Tn-C与Ca2+结合;
- Tn-T与原肌球蛋白有高度亲和力;
- Tn-I抑制肌球蛋白马达结构域的ATP酶活性;
肌肉收缩系统中的其他蛋白组分

(二)肌肉收缩的滑动模型
肌肉收缩的基本过程:
- 动作电位的产生;
- Ca2+的释放;
- 原肌球蛋白位移;
- 细肌丝和粗肌丝之间的相对滑动;
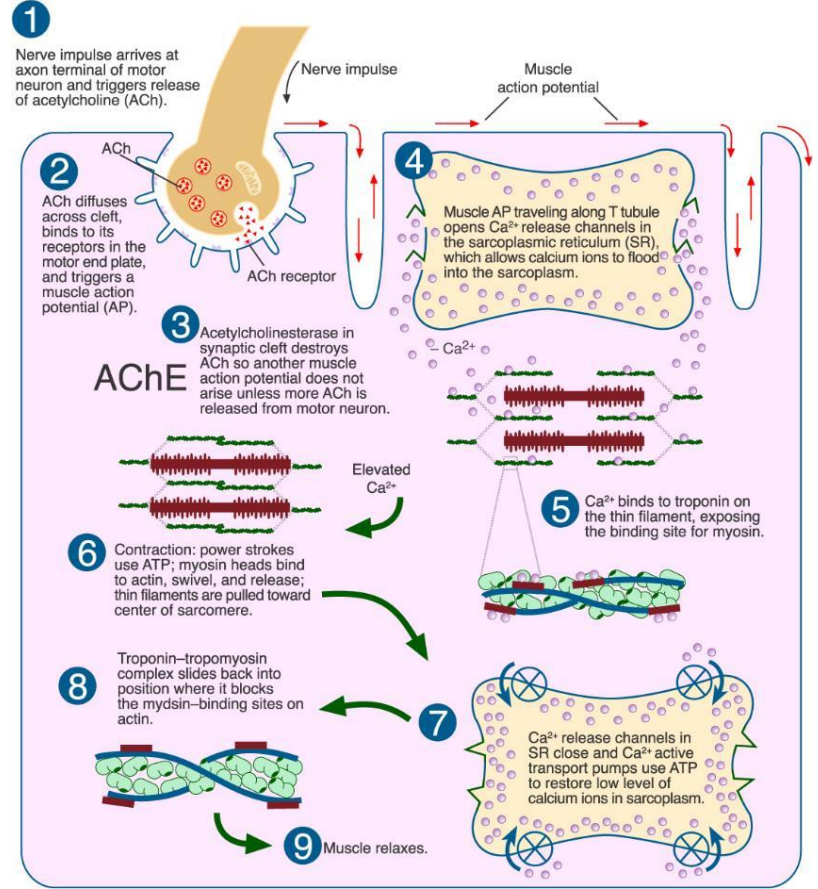
滑行学说(sliding theory)
Phosphorylation of myosin II by myosin light-chain kinase (MLCK)
第二节 微管及功能
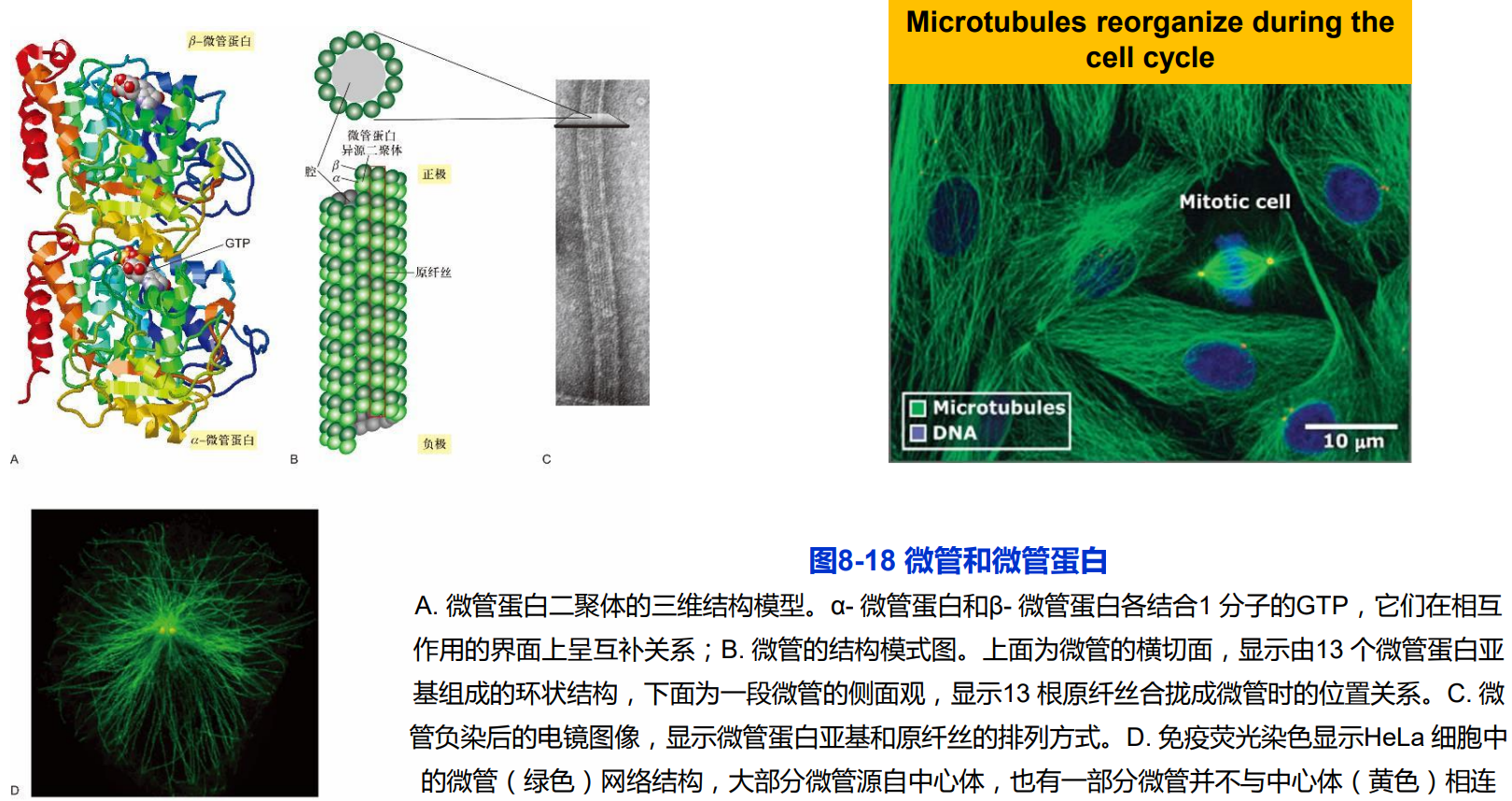
一、微管的结构组成与极性
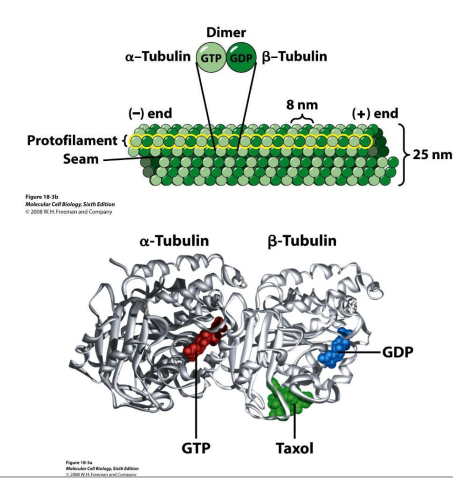
基本结构:α,β-微管蛋白二聚体;(细胞内游离态微管蛋白的主要存在形式)
GTP结合位点:
- 不可交换位点:α-微管蛋白;
- 可交换位点:β-微管蛋白;
二价阳离子结合位点;
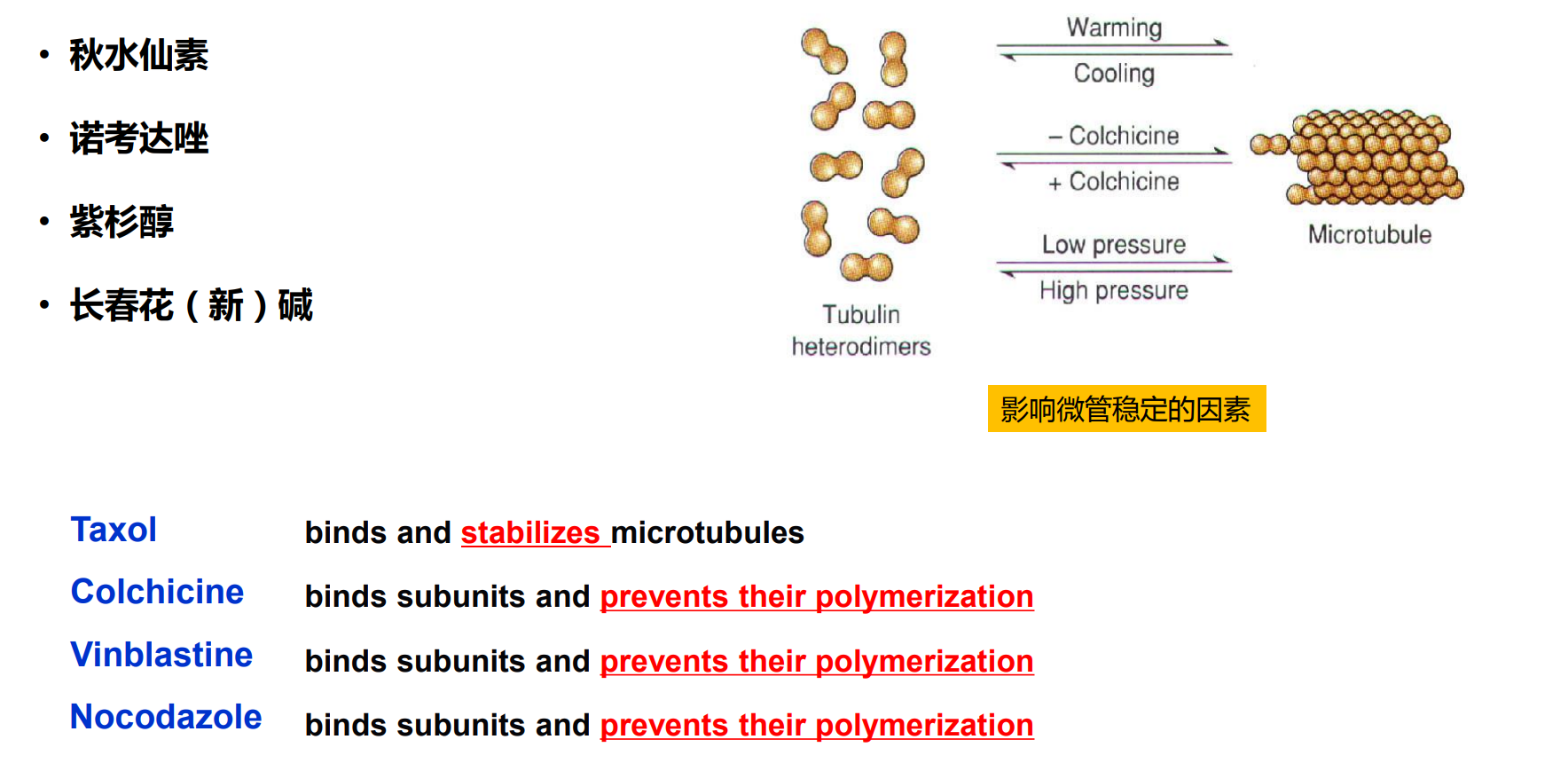
秋水仙素结合位点;
长春花碱结合位点;
13根原纤丝(protofilament)合拢形成微管的管壁;
极性:
- α-微管蛋白所在的一端为(-);
- β-微管蛋白所在一端为(+);
α/β-微管蛋白首先组装成原纤丝;原纤丝侧向相互作用形成片层;由13根原纤丝合拢形成的微管,α/β-微管蛋白从两端加入(或解聚)使微管延长(或缩短)。当体系中α/β-微管蛋白的浓度处于临界浓度时,微管蛋白在微管的正极端组装的速度与负极端去组装的速度相等,微管的长度可以保持不变。
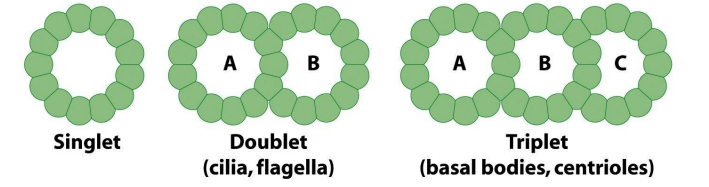
细胞内微管的3种类型
- 单管:细胞质微管或纺锤体微管;
- 二联管:纤毛或鞭毛中的轴丝微管;
- 三联管:中心体或基体中的微管;
二、微管的组装和去组装
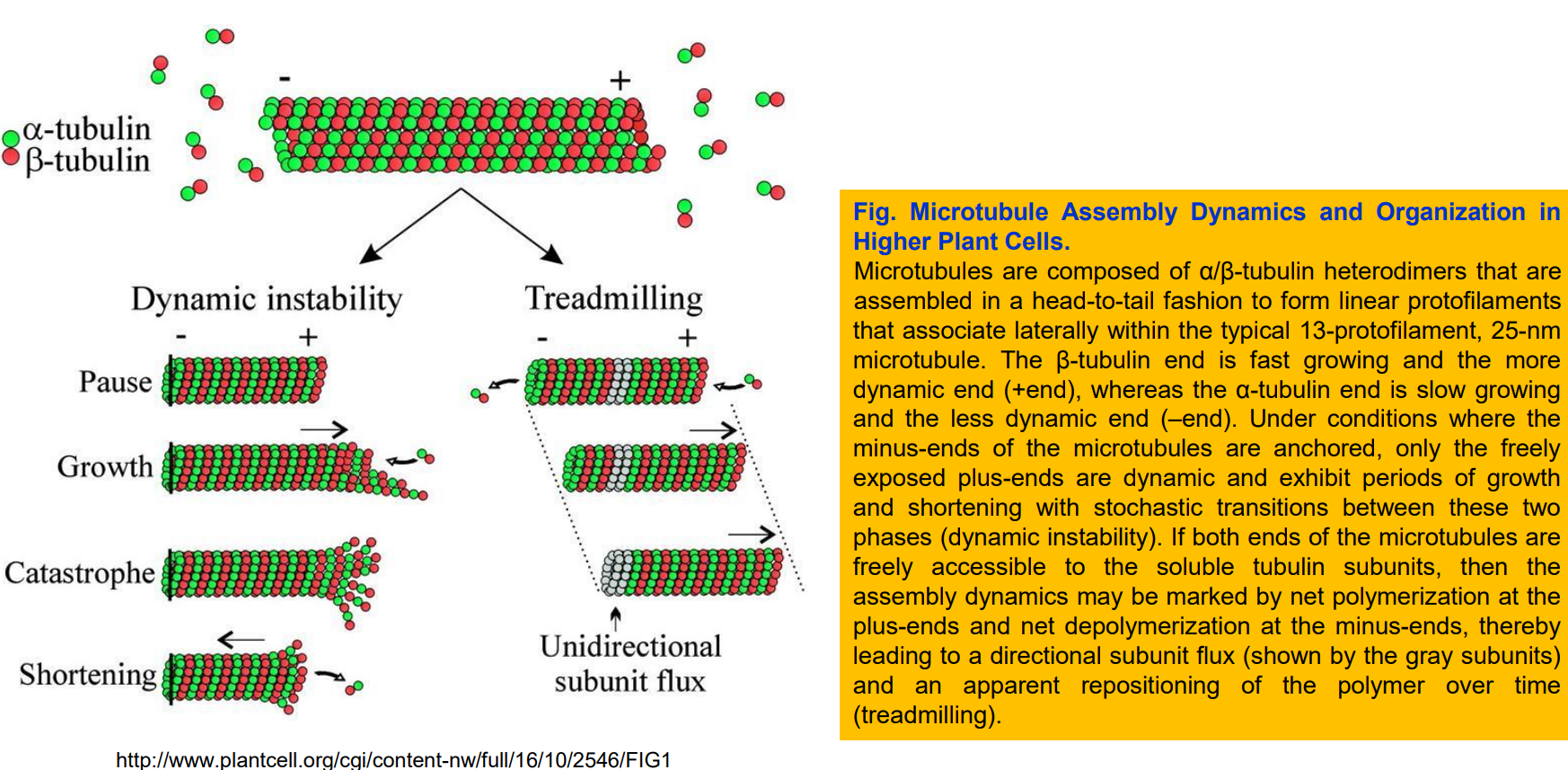
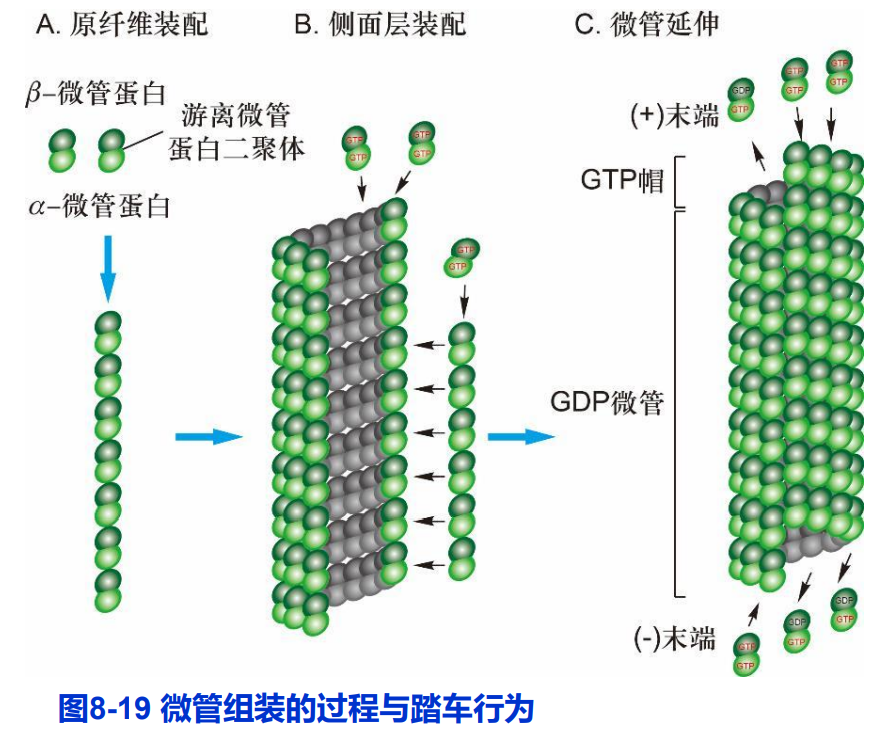
(一)微管的体外组装与踏车行为
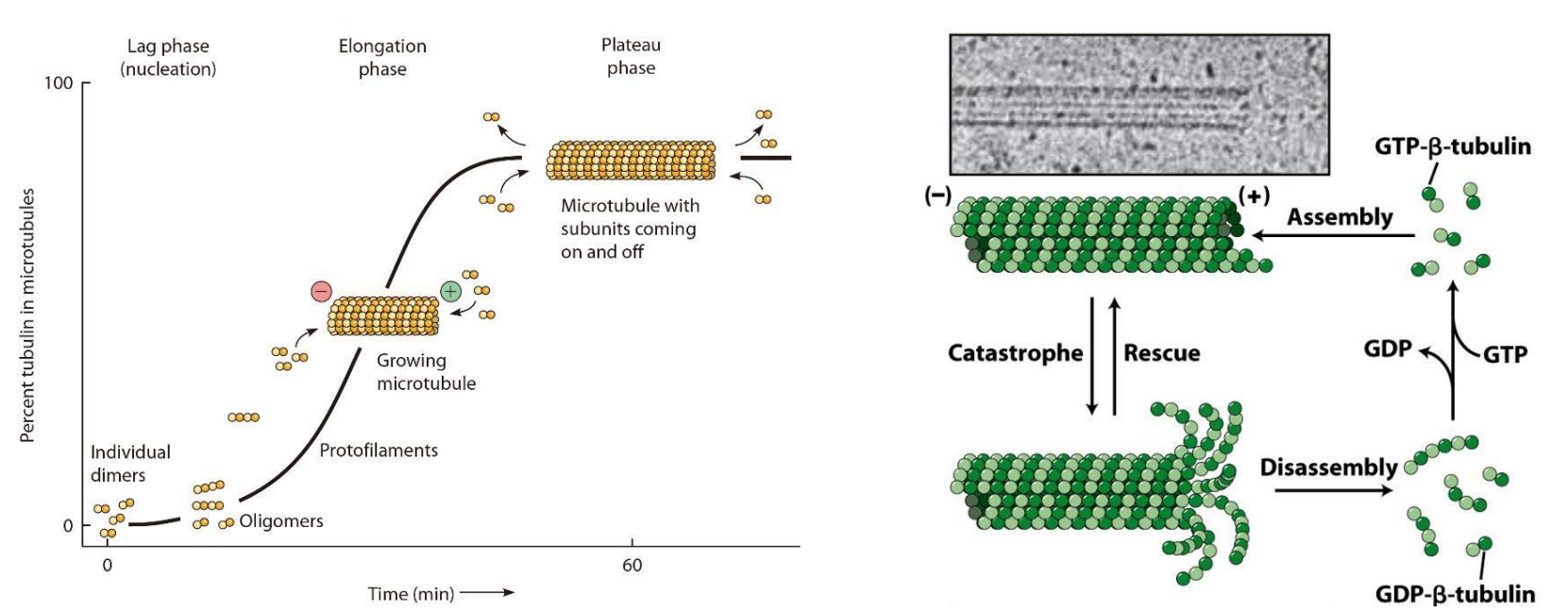
- 微管在体外组装的过程分为成核(nucleation)和延伸(elongation)两个阶段。
- 微管的组装速度同样与其底物(携带GTP的α/β-微管蛋白)的浓度正相关。
(二)作用于微管的特异性药物
三、微管组织中心(MTOC)体内组装
- MTOC(microtubule organizing centers):在细胞内起始微管的成核作用,并使之延伸的结构。
- 如动物细胞的细胞核附近的中心体,纤毛和鞭毛基部的基体以及上皮细胞顶端面高尔基体的反面网状结构。
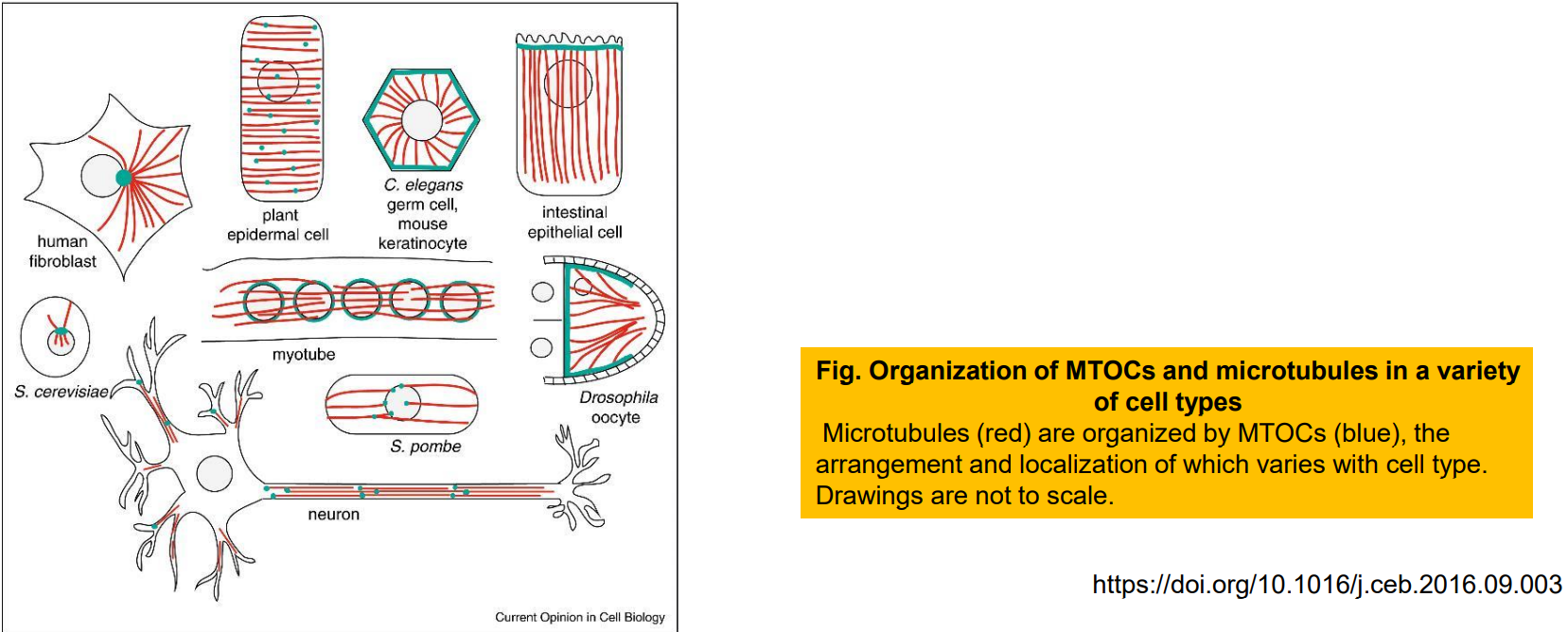
(一)中心体
- 中心体含有一对彼此垂直的桶装中心粒;
- 无定形的中心粒外周物质;9组三联体微管;
- 微管起源于中心粒外周物质区域(PCM);
- γ-微管蛋白在中心体的周质物质中形成的环状结构可诱导微管的成核和组装。
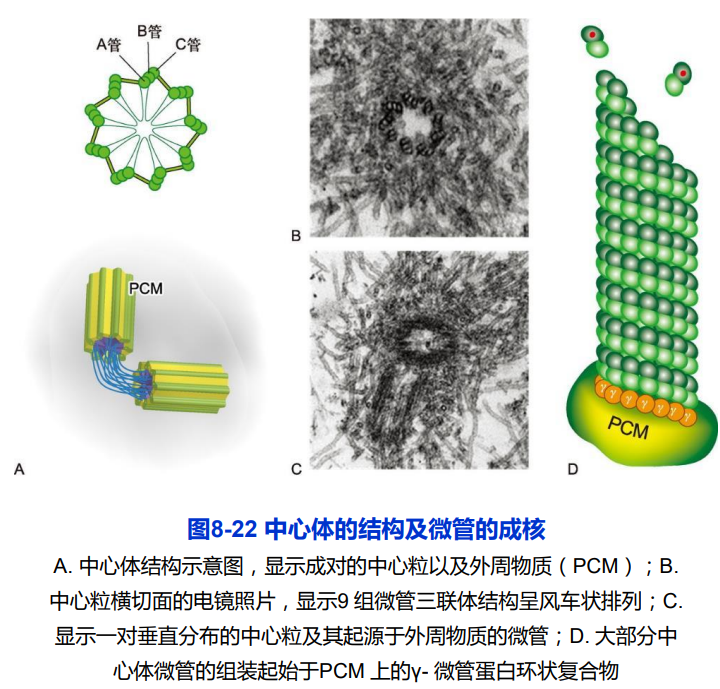
(二)基体和其他微管组织中心
- 基体:9组三联体微管;自我复制;
- 其他MTOC:高尔基体的反面膜囊区域和上皮细胞的顶端面等;
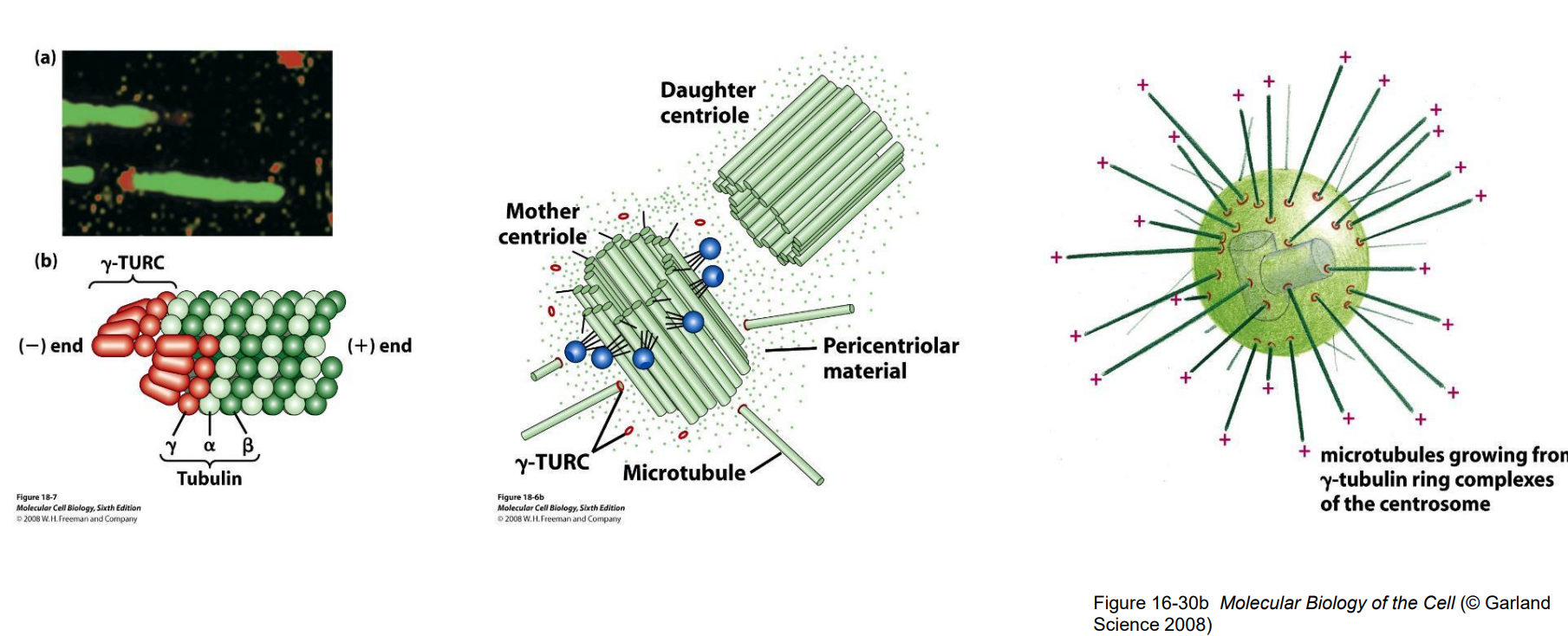
四、微管的动力学性质
- 微管的稳定性与其所结合的细胞结构组分以及细胞的生理状态相关
- 不同状态的微管其稳定性差异很大
- 在神经元分化过程中的作用
五、微管结合蛋白对微管网络结构的调节
- 微管结合蛋白(microtubule associated protein,MAP)
- 微管结合域(microtubule binding domain):与微管表面相互作用,稳定微管;
- 突出于微管表面结构域:与相邻微管或细胞结构相互作用,对微管网络的结构和功能进行调节。
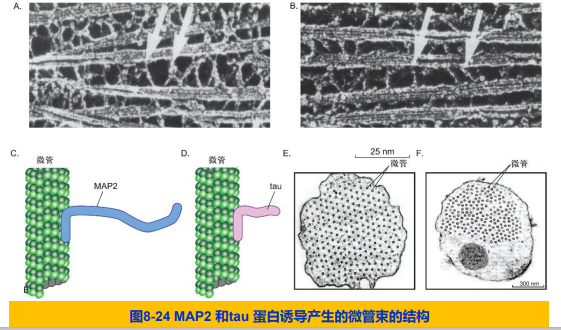

六、微管对细胞结构的组织作用
- 微管与细胞器的分布及细胞的形态发生和维持有很大关系;
- 微管极性排列方式与轴突物质运输密切相关;
- 物质沿着微管定向转移为细胞内各种细胞器和生物大分子的不对称分布提供了可能;
- 染色体运动;
七、细胞内依赖于微管的物质运输
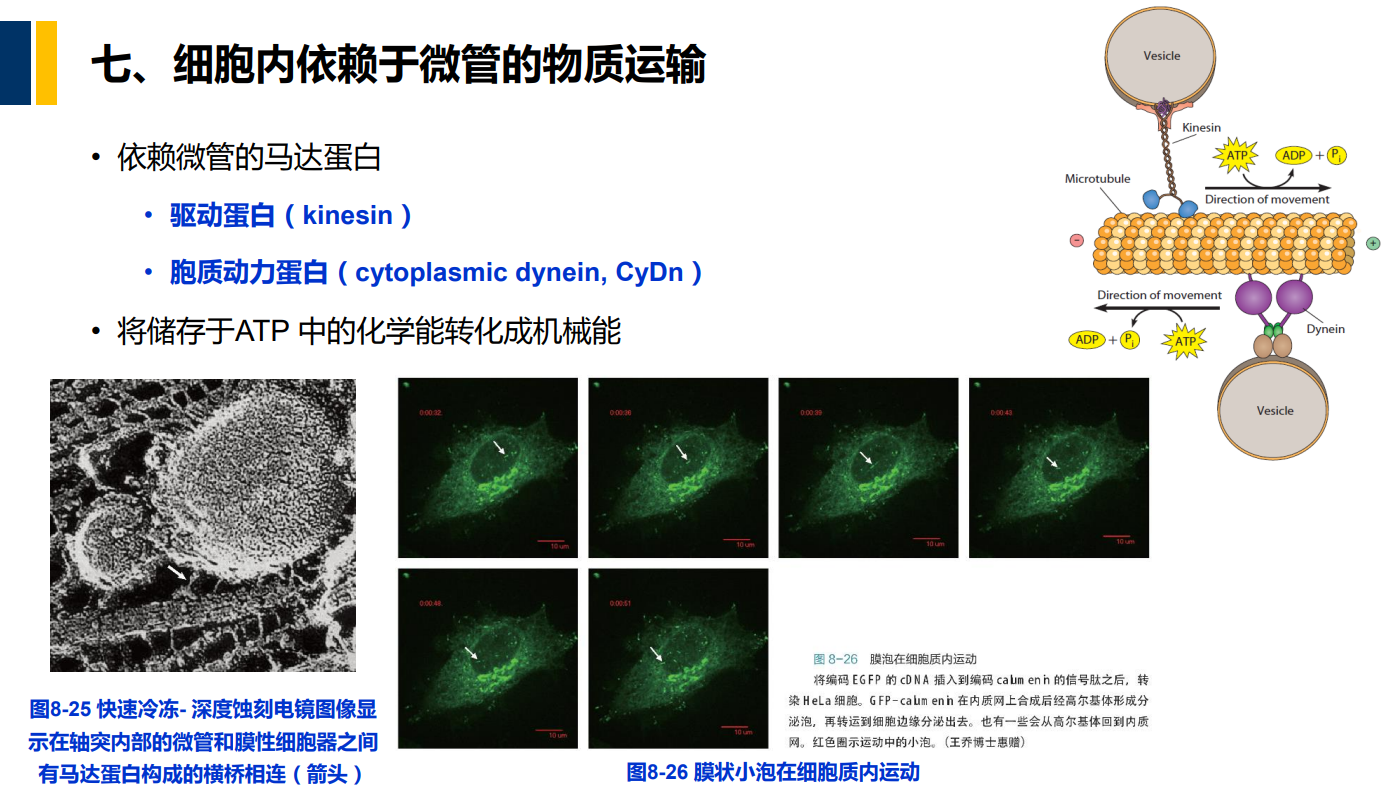
(一)驱动蛋白
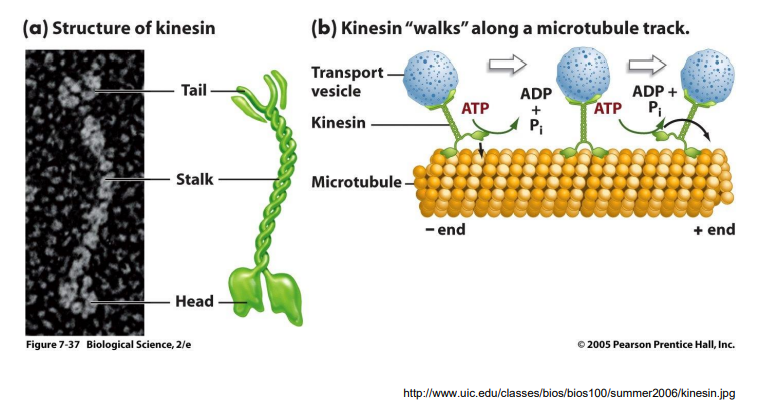
1. 驱动蛋白的分子结构及其功能
- 2条重链:具有马达结构域;
- 2条轻链:与重链尾部结合、具有货物结合功能;
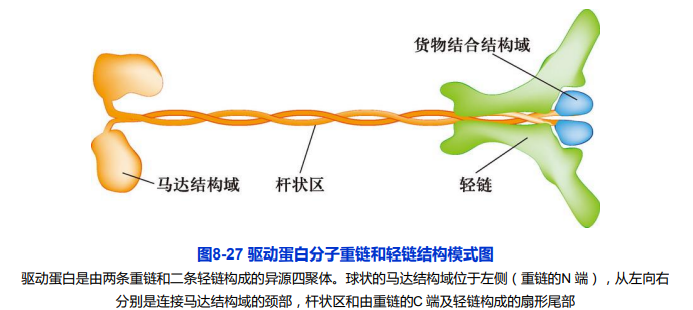
驱动蛋白家族成员的结构与功能
- 驱动蛋白超家族(kinesin superfamily protein,KIFs)
- N-驱动蛋白:马达结构域在肽链N-端,能从MT的负极向正极移动;
- M-驱动蛋白:马达结构域位于多肽链中部,结合在MT正极或负极端,使微管处于不稳定状态;
- C-驱动蛋白:马达结构域位于多肽链C-端,从微管的正极向负极移动;
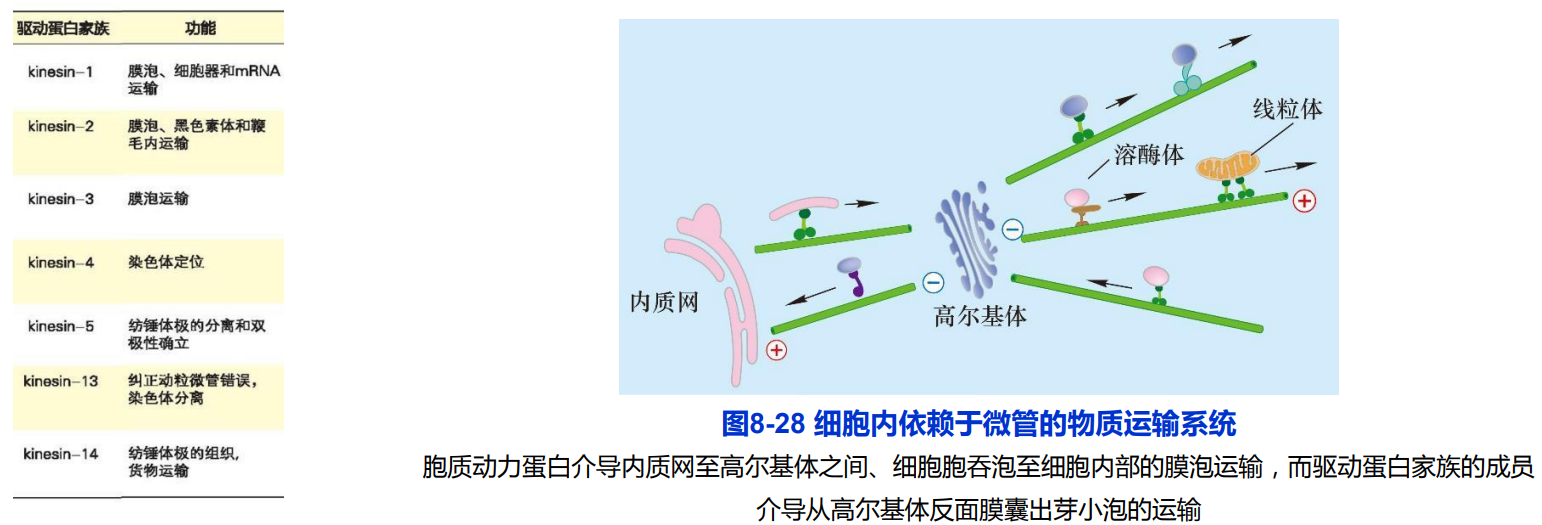
2. 驱动蛋白沿微管运动的分子机制
- 驱动蛋白的马达结构域上两个重要功能位点:ATP结合位点和微管结合位点;
- 驱动蛋白的运动主要涉及发生在两个马达结构域上ATP的结合、水解和ADP的释放以及与自身构象变化相偶联等机械化学循环过程。
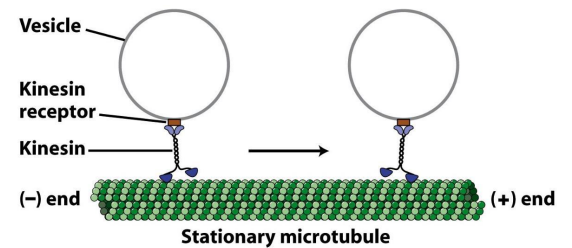
“步行” 模型和“尺蠖” 爬行模型
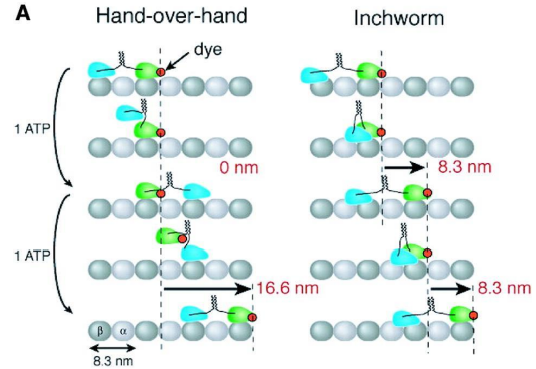
“步 行”(hand over hand) 模型
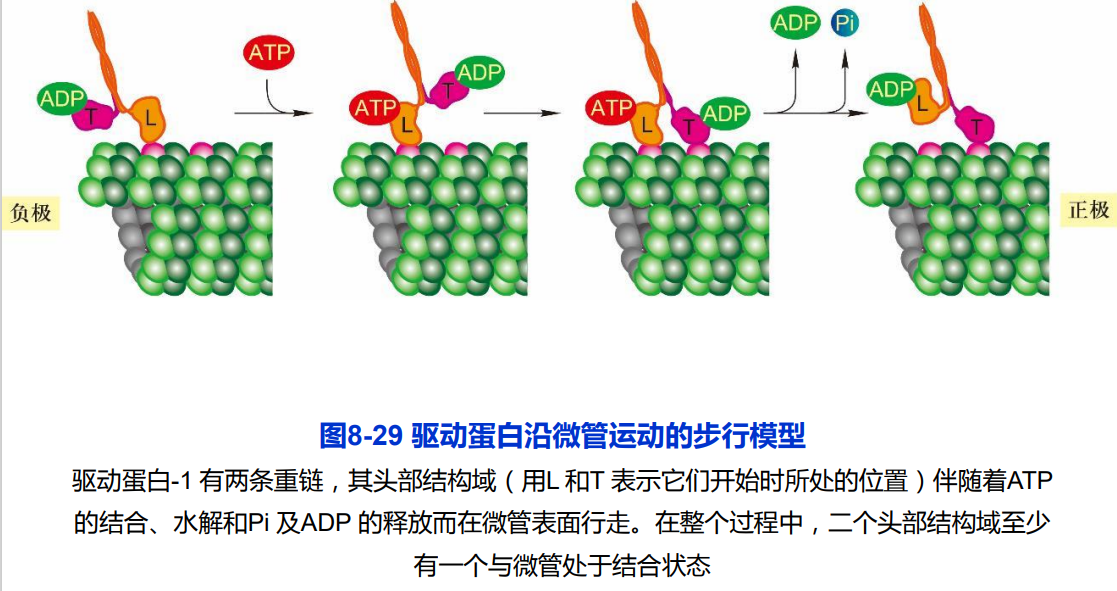
(二)胞质动力蛋白及其功能
动力蛋白超家族
- 细胞质动力蛋白(cytoplasmic dynein);
- 轴丝动力蛋白(axonemal dynein);
重链含ATP结合部位和微管结合部位;
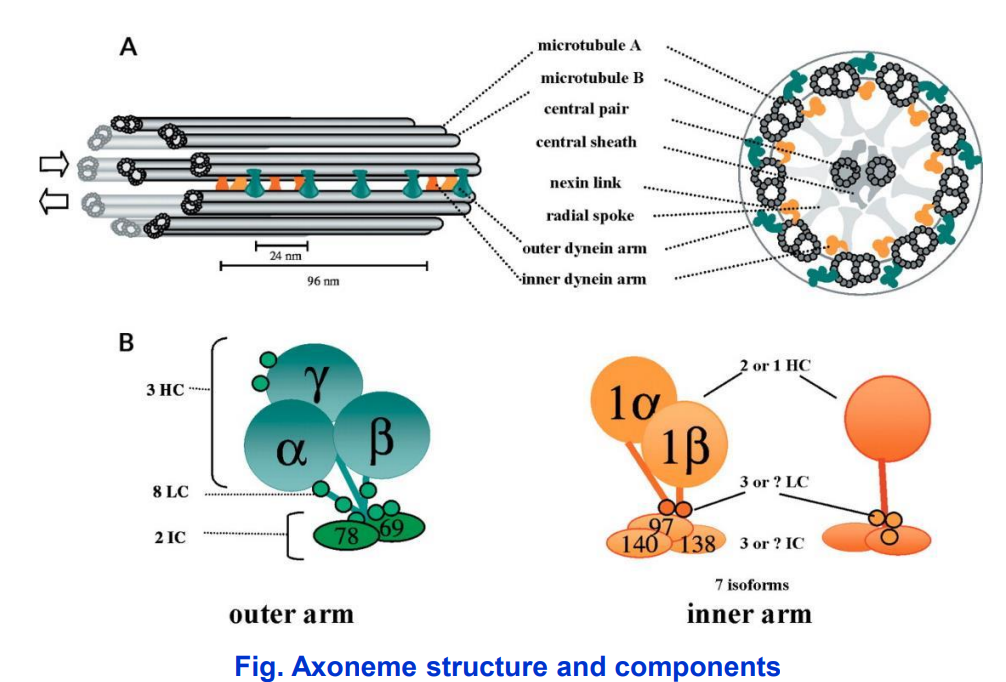
轴丝动力蛋白 axonemal dynein
• 内侧动力蛋白臂(inner dynein arms)和外侧动力蛋白臂(outer dynein arms)
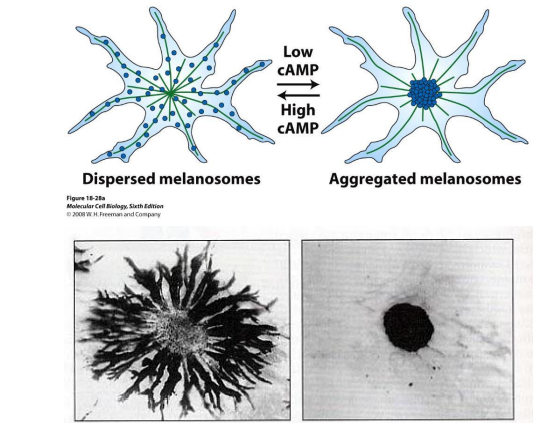
Movement of pigment granules: color adjustment
八、纤毛和鞭毛的结构与功能
纤毛(cilia)和鞭毛(flagella)是突出于细胞表面的、高度特化的细胞结构。
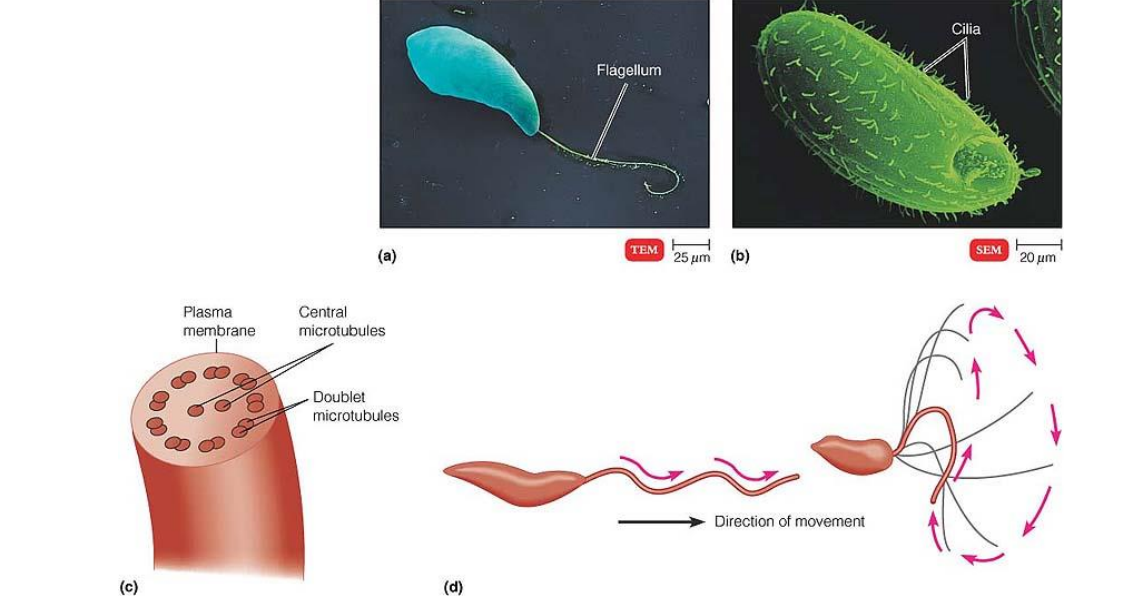
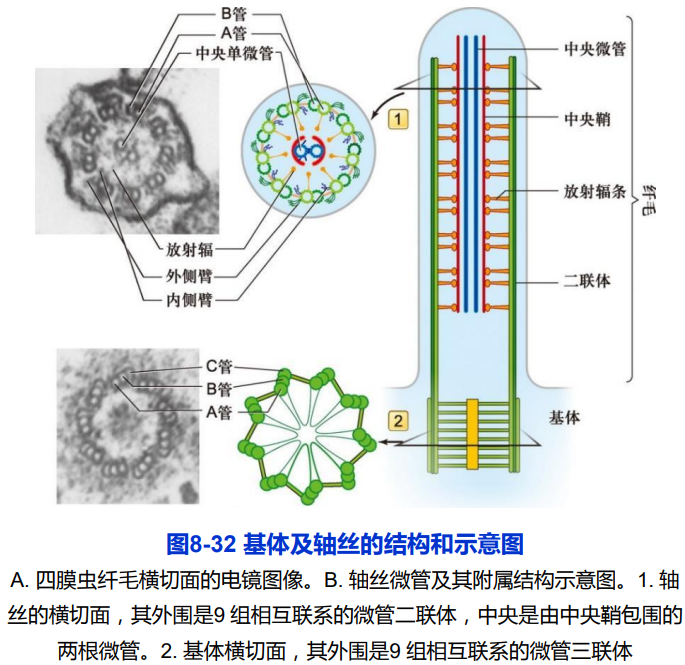
(一)纤毛的结构及组装
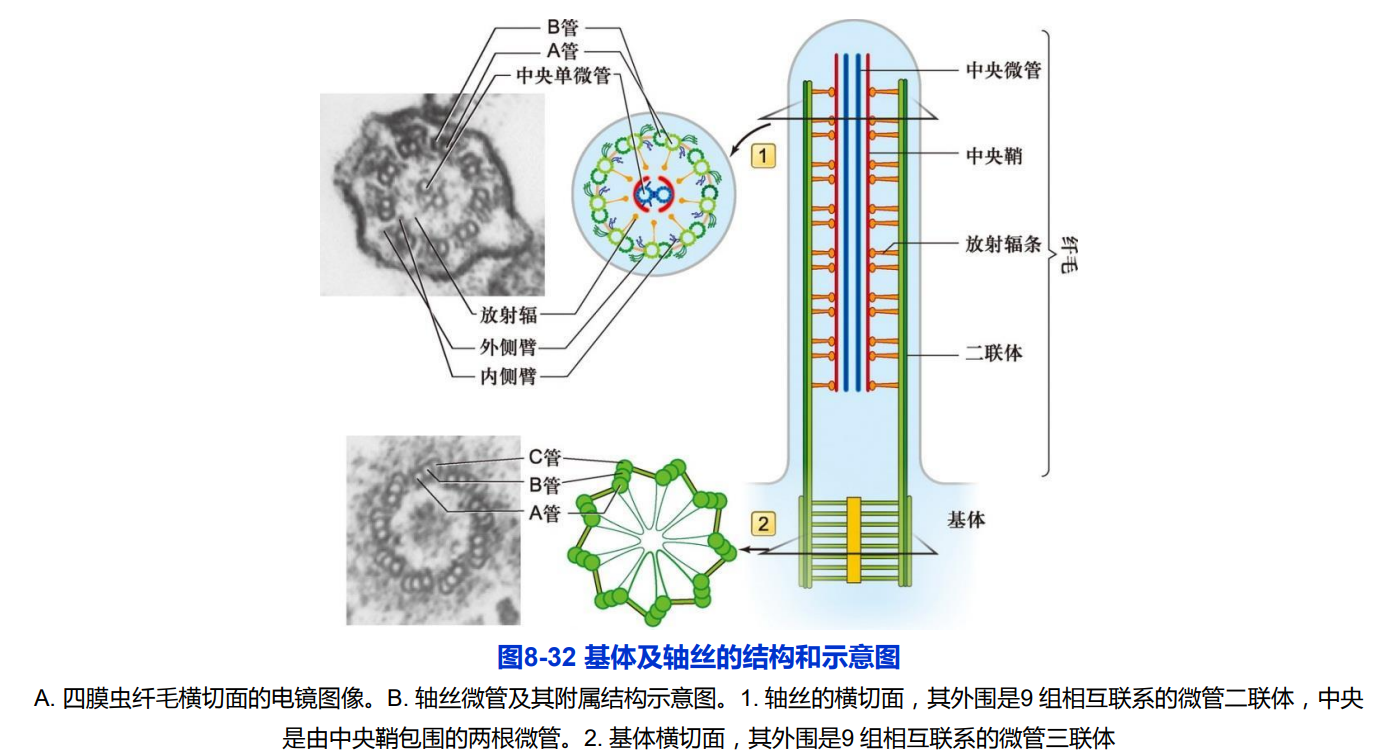
1. 纤毛的结构
- 纤毛的外部是由细胞质膜的特化而形成的纤毛膜,内部是由微管及其附属蛋白组装而成的轴丝;
- 轴丝是由250多种不同的蛋白质组装而成的高度有序的结构;
- 轴丝微管排列方式主要有三种模式:9+2;9+0;9+4;
- 轴丝微管正极端都有指向纤毛或鞭毛顶端。外围二联体微管由A管和B管组成,A管为完全微管,由13个亚基环绕而成,B管为不完全微管,仅由10个亚基构成。中央微管均为完全微管。
- 基体外围含有9组三联体微管,没有中央微管,呈“9+0”排列
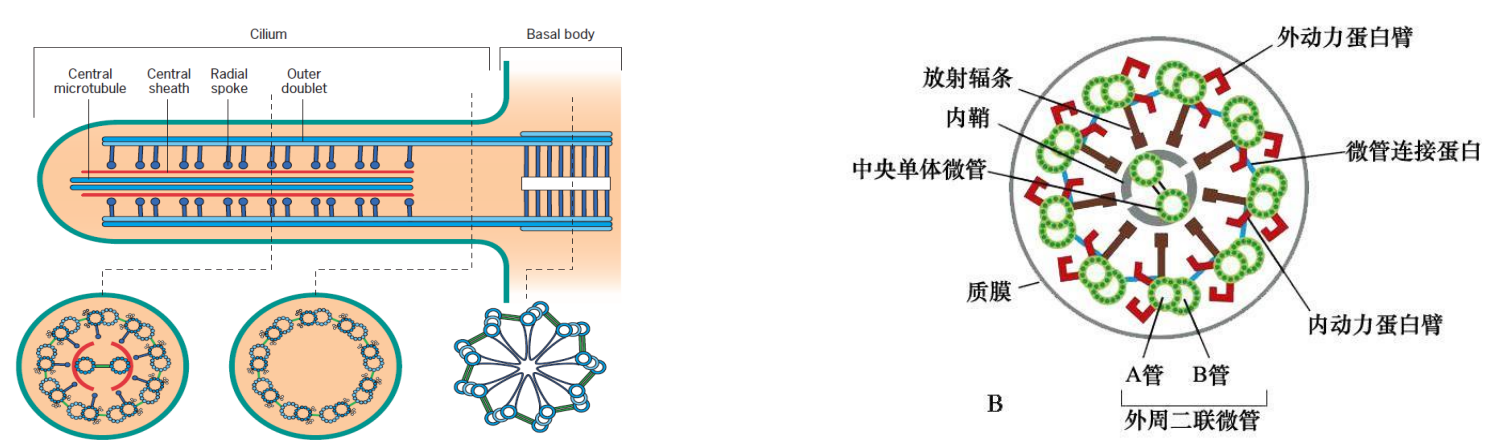
2. 纤毛的组装(发生)
- 起始于基体;
- 原生纤毛的组成和细胞周期紧密相关;
- 原生纤毛的形成分为四个阶段;
- 纤毛的延伸和维持依赖于鞭毛内运输。
(二)纤毛或鞭毛的运动机制
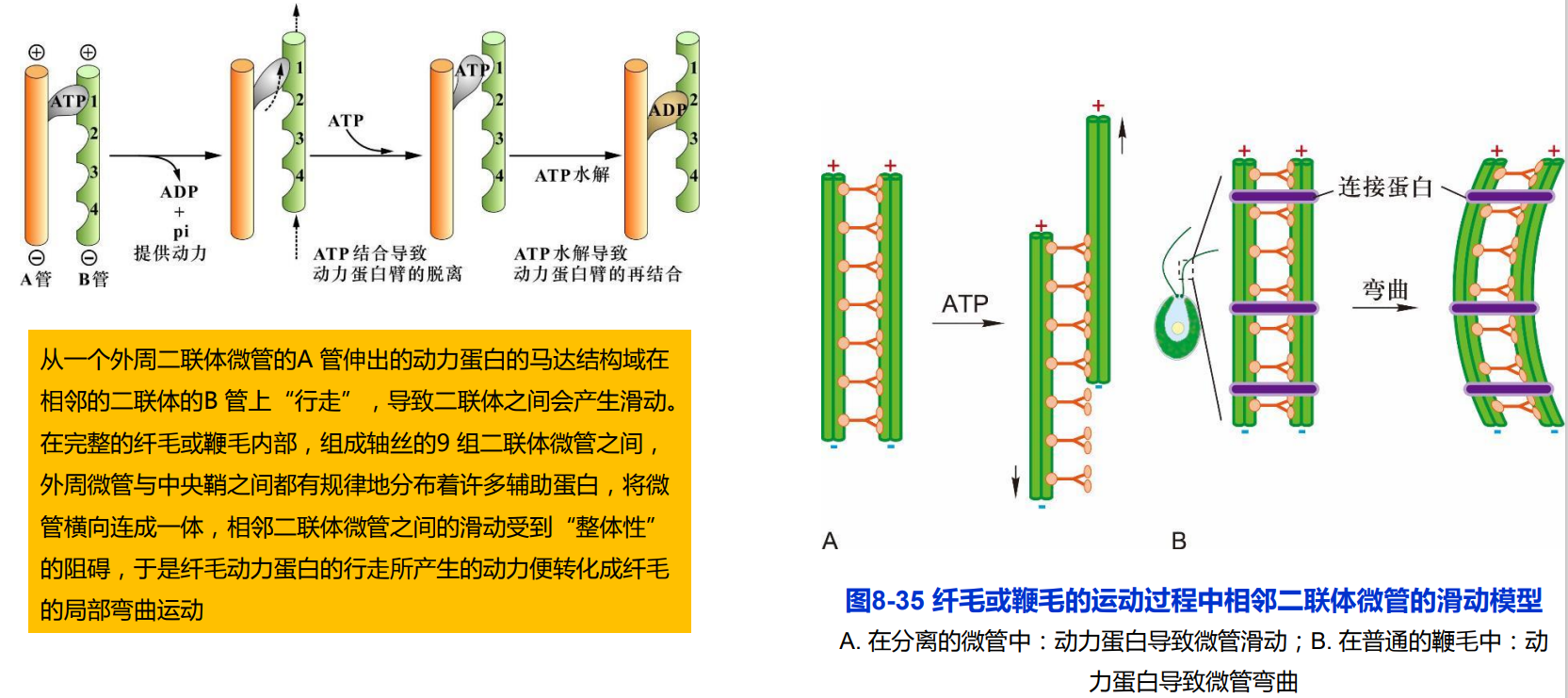
细菌鞭毛运动机制
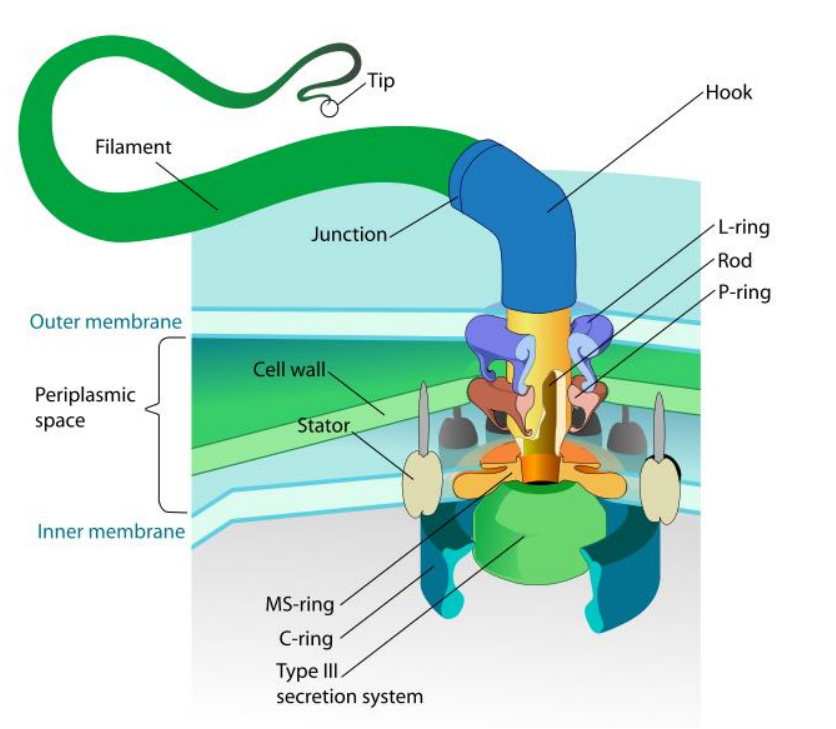
(三)纤毛的功能
运动装置;
物理感受器(mechanosensor);
化学感受器;
动物胚胎发育过程中,影响躯体各器官正常分布;
参与了发育过程中两类信号通路:
- Hh信号通路;
- Wnt信号通路;
九、纺锤体和染色体运动
纺锤体微管包括动粒微管、极微管和星体微管。
- 动粒微管连接染色体动粒与中心体。
- 极微管从两极发出,在纺锤体中部赤道区相互交错重叠。
- 星体微管从中心体向周围呈辐射状分布
有丝分裂过程中染色体的运动有赖于纺锤体微管的组装和解离。
第三节 中间丝
中间丝又称中间纤维(intermediate filament,IF)
最初是在平滑肌细胞内发现;直径10 nm ;粗细介于肌细胞的粗肌丝和细肌丝之间;存在于绝大多数动物细胞
一、中间丝的主要类型和组成成分
- 不同组织来源的细胞表达不同类型的中间丝蛋白
- 根据中间丝蛋白的氨基酸序列、基因结构、组装特性以及在发育过程的组织特异性表达模式等, 可分为6 种主要类型

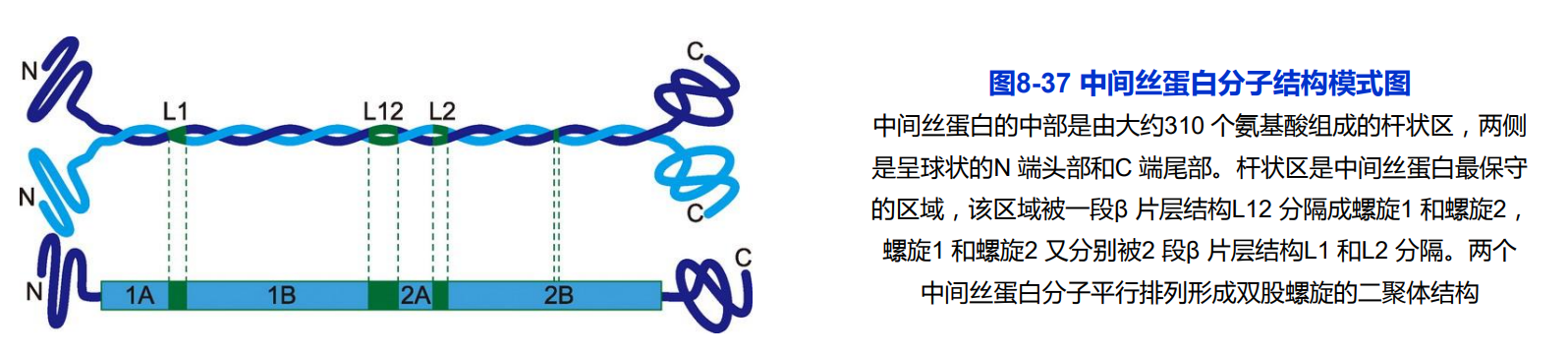
中间丝蛋白分子结构
- 不同种类的中间丝蛋白有非常相似的二级结构;
- 高度保守的杆状区;
- 高度多变的头部和尾部;
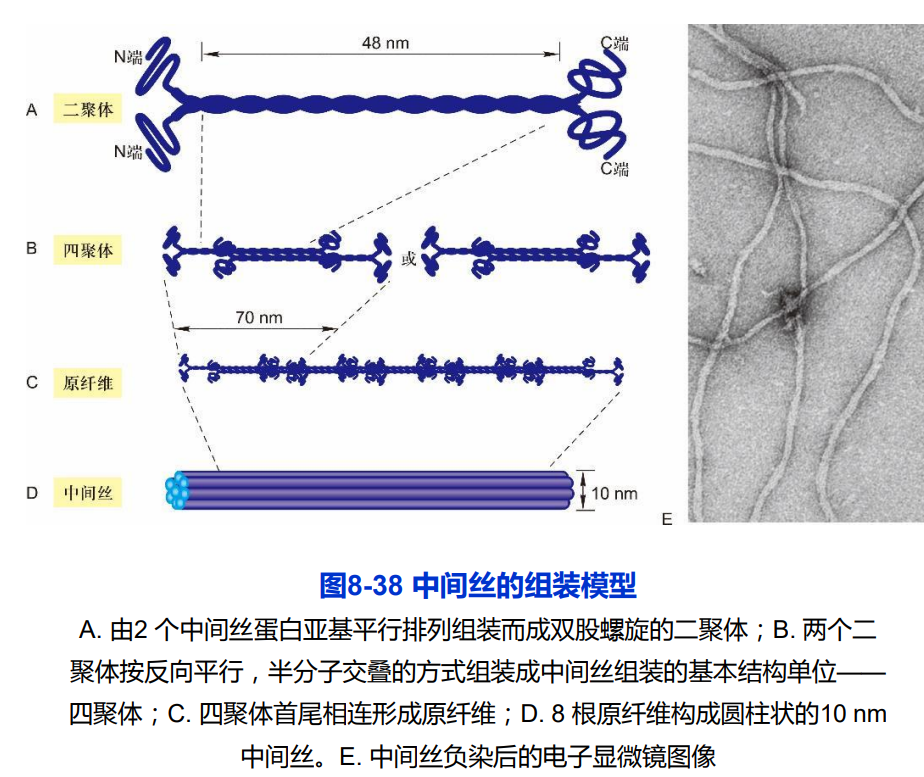
二、中间丝的组装与表达
- IF没有极性
- 不需要ATP、GTP参与
- 不表现为典型的踏车行为
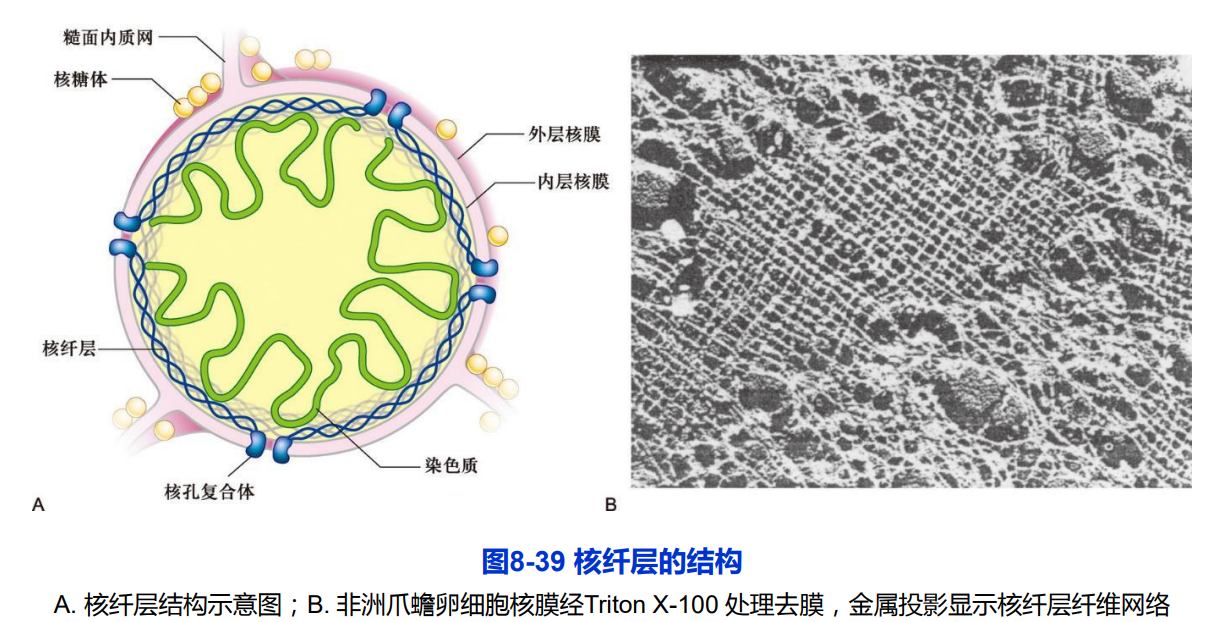
三、中间丝与其他细胞结构的联系
• 核纤层:磷酸化与去磷酸化
中间丝结合蛋白

中间丝的功能
- 提供机械稳定性;
- 维持细胞核膜稳定;
- 参与细胞连接;
- 参与信号传导;
- 参与细胞分化;
单纯性疱性表皮松懈症(simplex epidermolysis bullosa)
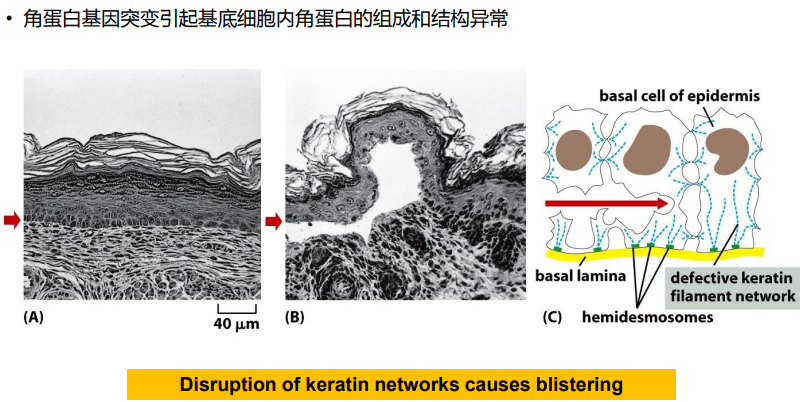
本章内容提要
,具有组织和表达阶段的特异性。存在ATP/ADP结合位点;
α- 肌动蛋白为横纹肌、心肌、血管平滑肌和肠道平滑肌所特有;
β- 肌动蛋白和 γ-肌动蛋白存在于所有的细胞中
ATP帽(ATP Cap)与踏车行为(treadmilling)
当微丝末端聚合速度大于ATP水解速度时,会在末端形成一个ATP帽;
踏车行为:在体外组装过程中,微丝正极由于肌动蛋白亚基不断增长而延长,负极由于肌动蛋白亚基去组装而缩短的现象。
微丝细胞内组装:受微丝结合蛋白影响
(三)影响微丝组装的特异性药物
细胞松弛素(cytochalasin)
- 与微丝结合之后将微丝切断,并结合在微丝末端阻碍肌动蛋白的聚合,但对微丝解聚没有影响;
- 破坏微丝网络结构,并阻止细胞的运动。
鬼笔环肽(phalloidin)
- 与微丝表面有强亲和力,不与肌动蛋白单体结合;
- 阻止微丝的解聚,使其保持稳定状态。
二、微丝网络结构的调节与细胞运动
细胞中大多数微丝结构处于动态的组装和去组装过程,并通过这种方式实现功能。
(一)非肌肉细胞内微丝的结合蛋白
1.肌动蛋白单体结合蛋白
结合ATP的肌动蛋白既可以参与微丝正极端的组装,也可以在负极端进行组装。胸腺素β4与肌动蛋白结合以后抑制微丝的组装。前纤维蛋白与肌动蛋白单体的底部结合,促进了微丝正极端的组装,但阻断了负极端的组装。
2. 成核蛋白-Arp2/3促进微丝蛋白的分叉
3. 加帽蛋白(capping protein)-增加稳定性
4. 交联蛋白
成束蛋白(bunding protein):将相邻的微丝交联成平行排列状态;
凝胶形成蛋白(gel-forming protein):将微丝联接成网状。
5. 割断及解聚蛋白
凝溶胶蛋白(gelsolin):在高Ca2+(>1 μmol/L)情况下 能将较长微丝切断,使肌动蛋白由凝胶态转化成溶胶态
丝切蛋白/ 肌动蛋白解聚因子(cofilin/actin depolymerizing factor,Cofilin/ADF):能与游离的肌动 蛋白或微丝结合,提高微丝的解聚速度
(二)细胞皮层(cell cortex)
细胞内大部分微丝都集中在紧贴细胞质膜的细胞质区域,并由微丝交联蛋白交联成凝胶态的三维网络结构。
限制膜蛋白的流动性;为细胞质膜提供强度和韧性;维持细胞形状;细胞的多种运动,如胞质 环流(cyclosis)、阿米巴运动(amoiboid)、变皱膜运动 (ruffled membrane locomotion)、 吞噬(phagocytosis)以及膜蛋白的定位等与皮层内肌动蛋白的溶胶态- 凝胶态转化相关。
(三)应力纤维
(四)细胞伪足的形成与细胞迁移
三、肌球蛋白:依赖于微丝的分子马达
马达蛋白可分为三种:
- 沿微丝运动的肌球蛋白(myosin);
- 沿微管移动的驱动蛋白(kinesin);
- 沿微管移动的动力蛋白(dynein);
马达结构域与微丝或微管结合;货物结合结构域与膜性细胞器或大分子复合物特异性结合;
能量转换:利用水解ATP所提供的能量沿微丝或微管运动。
(一)肌球蛋白的种类
- 骨骼肌细胞内,多个Ⅱ型肌球蛋白分子组装成肌原纤维的粗肌丝;
- 马达结构域是肌球蛋白超家族成员最保守的部位,是肌球蛋白定性和分类的依据;
- 基于马达结构域多肽链一级结构的相似性,至少可以将肌球蛋白超家族成员分成18种家族;
(二)肌球蛋白的结构
肌球蛋白是沿着微丝运动的马达蛋白,通常有三个功能结构域;
- 马达结构域:位于肌球蛋白的头部,包含一个肌动蛋白亚基结合位点和一个具有ATP酶活性的ATP结合位点,负责将ATP水解释放发化学能转变为机械能。
- 调控结构域:连接马达结构域和尾部杆状区的一段α螺旋,轻链的结合部位,发挥杠杆作用。
- 尾部结构域:选择性与所运输的“货物”结合;
1. Ⅱ型肌球蛋白:传统的肌球蛋白(conventional myosin)
- 肌细胞:组装成肌原纤维的粗丝;
- 非肌细胞:胞质分裂过程中收缩环的主要结构成分;应力纤维的结构成分;
- 包含两条重链和4条轻链,形成一个高度不对称的结构。
2. 其他类型的肌球蛋白(unconventional myosin)
- Ⅰ型肌球蛋白分子:一个头部(马达结构域)和一个尾部
- Ⅴ型肌球蛋白分子:两条肽链组成的二聚体;两个头部交替与微丝结合
四、肌细胞的收缩运动
(一)肌纤维的结构
- 肌纤维:肌原纤维(myofibrils);
- 肌节(sarcomere);
- 粗肌丝;细肌丝;
- 横桥;
细肌丝结构
肌动蛋白;
- 原肌球蛋白(tropomyosin,Tm);
- 两条平行多肽形成α螺旋构型,调节actin与myosin头部结合;
肌钙蛋白(troponin,Tn):
Tn-C与Ca2+结合;
- Tn-T与原肌球蛋白有高度亲和力;
- Tn-I抑制肌球蛋白马达结构域的ATP酶活性;
肌肉收缩系统中的其他蛋白组分
(二)肌肉收缩的滑动模型
肌肉收缩的基本过程:
- 动作电位的产生;
- Ca2+的释放;
- 原肌球蛋白位移;
- 细肌丝和粗肌丝之间的相对滑动;
滑行学说(sliding theory)
Phosphorylation of myosin II by myosin light-chain kinase (MLCK)
第二节 微管及功能
一、微管的结构组成与极性
基本结构:α,β-微管蛋白二聚体;(细胞内游离态微管蛋白的主要存在形式)
GTP结合位点:
- 不可交换位点:α-微管蛋白;
- 可交换位点:β-微管蛋白;
二价阳离子结合位点;
秋水仙素结合位点;
长春花碱结合位点;
13根原纤丝(protofilament)合拢形成微管的管壁;
极性:
- α-微管蛋白所在的一端为(-);
- β-微管蛋白所在一端为(+);
α/β-微管蛋白首先组装成原纤丝;原纤丝侧向相互作用形成片层;由13根原纤丝合拢形成的微管,α/β-微管蛋白从两端加入(或解聚)使微管延长(或缩短)。当体系中α/β-微管蛋白的浓度处于临界浓度时,微管蛋白在微管的正极端组装的速度与负极端去组装的速度相等,微管的长度可以保持不变。
细胞内微管的3种类型
- 单管:细胞质微管或纺锤体微管;
- 二联管:纤毛或鞭毛中的轴丝微管;
- 三联管:中心体或基体中的微管;
二、微管的组装和去组装
(一)微管的体外组装与踏车行为
- 微管在体外组装的过程分为成核(nucleation)和延伸(elongation)两个阶段。
- 微管的组装速度同样与其底物(携带GTP的α/β-微管蛋白)的浓度正相关。
(二)作用于微管的特异性药物
三、微管组织中心(MTOC)体内组装
- MTOC(microtubule organizing centers):在细胞内起始微管的成核作用,并使之延伸的结构。
- 如动物细胞的细胞核附近的中心体,纤毛和鞭毛基部的基体以及上皮细胞顶端面高尔基体的反面网状结构。
(一)中心体
- 中心体含有一对彼此垂直的桶装中心粒;
- 无定形的中心粒外周物质;9组三联体微管;
- 微管起源于中心粒外周物质区域(PCM);
- γ-微管蛋白在中心体的周质物质中形成的环状结构可诱导微管的成核和组装。
(二)基体和其他微管组织中心
- 基体:9组三联体微管;自我复制;
- 其他MTOC:高尔基体的反面膜囊区域和上皮细胞的顶端面等;
四、微管的动力学性质
- 微管的稳定性与其所结合的细胞结构组分以及细胞的生理状态相关
- 不同状态的微管其稳定性差异很大
- 在神经元分化过程中的作用
五、微管结合蛋白对微管网络结构的调节
- 微管结合蛋白(microtubule associated protein,MAP)
- 微管结合域(microtubule binding domain):与微管表面相互作用,稳定微管;
- 突出于微管表面结构域:与相邻微管或细胞结构相互作用,对微管网络的结构和功能进行调节。
六、微管对细胞结构的组织作用
- 微管与细胞器的分布及细胞的形态发生和维持有很大关系;
- 微管极性排列方式与轴突物质运输密切相关;
- 物质沿着微管定向转移为细胞内各种细胞器和生物大分子的不对称分布提供了可能;
- 染色体运动;
七、细胞内依赖于微管的物质运输
(一)驱动蛋白
1. 驱动蛋白的分子结构及其功能
- 2条重链:具有马达结构域;
- 2条轻链:与重链尾部结合、具有货物结合功能;
驱动蛋白家族成员的结构与功能
- 驱动蛋白超家族(kinesin superfamily protein,KIFs)
- N-驱动蛋白:马达结构域在肽链N-端,能从MT的负极向正极移动;
- M-驱动蛋白:马达结构域位于多肽链中部,结合在MT正极或负极端,使微管处于不稳定状态;
- C-驱动蛋白:马达结构域位于多肽链C-端,从微管的正极向负极移动;
2. 驱动蛋白沿微管运动的分子机制
- 驱动蛋白的马达结构域上两个重要功能位点:ATP结合位点和微管结合位点;
- 驱动蛋白的运动主要涉及发生在两个马达结构域上ATP的结合、水解和ADP的释放以及与自身构象变化相偶联等机械化学循环过程。
“步行” 模型和“尺蠖” 爬行模型
“步 行”(hand over hand) 模型
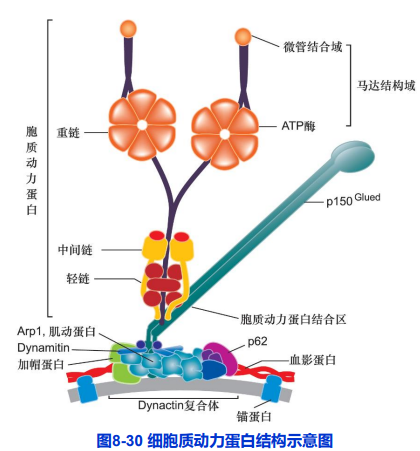
(二)胞质动力蛋白及其功能
动力蛋白超家族
- 细胞质动力蛋白(cytoplasmic dynein);
- 轴丝动力蛋白(axonemal dynein);
重链含ATP结合部位和微管结合部位;
轴丝动力蛋白 axonemal dynein
• 内侧动力蛋白臂(inner dynein arms)和外侧动力蛋白臂(outer dynein arms)
Movement of pigment granules: color adjustment
八、纤毛和鞭毛的结构与功能
纤毛(cilia)和鞭毛(flagella)是突出于细胞表面的、高度特化的细胞结构。
(一)纤毛的结构及组装
1. 纤毛的结构
- 纤毛的外部是由细胞质膜的特化而形成的纤毛膜,内部是由微管及其附属蛋白组装而成的轴丝;
- 轴丝是由250多种不同的蛋白质组装而成的高度有序的结构;
- 轴丝微管排列方式主要有三种模式:9+2;9+0;9+4;
- 轴丝微管正极端都有指向纤毛或鞭毛顶端。外围二联体微管由A管和B管组成,A管为完全微管,由13个亚基环绕而成,B管为不完全微管,仅由10个亚基构成。中央微管均为完全微管。
- 基体外围含有9组三联体微管,没有中央微管,呈“9+0”排列
2. 纤毛的组装(发生)
- 起始于基体;
- 原生纤毛的组成和细胞周期紧密相关;
- 原生纤毛的形成分为四个阶段;
- 纤毛的延伸和维持依赖于鞭毛内运输。
(二)纤毛或鞭毛的运动机制
细菌鞭毛运动机制
(三)纤毛的功能
运动装置;
物理感受器(mechanosensor);
化学感受器;
动物胚胎发育过程中,影响躯体各器官正常分布;
参与了发育过程中两类信号通路:
- Hh信号通路;
- Wnt信号通路;
九、纺锤体和染色体运动
纺锤体微管包括动粒微管、极微管和星体微管。
- 动粒微管连接染色体动粒与中心体。
- 极微管从两极发出,在纺锤体中部赤道区相互交错重叠。
- 星体微管从中心体向周围呈辐射状分布
有丝分裂过程中染色体的运动有赖于纺锤体微管的组装和解离。
第三节 中间丝
中间丝又称中间纤维(intermediate filament,IF)
最初是在平滑肌细胞内发现;直径10 nm ;粗细介于肌细胞的粗肌丝和细肌丝之间;存在于绝 大多数动物细胞
一、中间丝的主要类型和组成成分
- 不同组织来源的细胞表达不同类型的中间丝蛋白
- 根据中间丝蛋白的氨基酸序列、基因结构、组装特性以及在发育过程的组织特异性表达模式等, 可分为6 种主要类型
中间丝蛋白分子结构
- 不同种类的中间丝蛋白有非常相似的二级结构;
- 高度保守的杆状区;
- 高度多变的头部和尾部;
二、中间丝的组装与表达
- IF没有极性
- 不需要ATP、GTP参与
- 不表现为典型的踏车行为
三、中间丝与其他细胞结构的联系
• 核纤层:磷酸化与去磷酸化
中间丝结合蛋白
中间丝的功能
- 提供机械稳定性;
- 维持细胞核膜稳定;
- 参与细胞连接;
- 参与信号传导;
- 参与细胞分化;
单纯性疱性表皮松懈症(simplex epidermolysis bullosa)
本章内容提要
最后
以上就是玩命魔镜最近收集整理的关于细胞生物学8-第八章-细胞骨架0.细胞骨架的基本概念第一节 微丝与细胞运动0.细胞骨架的基本概念第一节 微丝与细胞运动 第二节 微管及功能第三节 中间丝 本章内容提要 第二节 微管及功能第三节 中间丝 本章内容提要的全部内容,更多相关细胞生物学8-第八章-细胞骨架0.细胞骨架的基本概念第一节内容请搜索靠谱客的其他文章。








发表评论 取消回复