文章目录
- 传统甲烷氧化菌主导稻田土壤中大气甲烷的氧化
- 写在前面
- 摘要
- 背景
- 结果与讨论
- 土壤中HAMO活性的发生与恢复
- 图1 水稻土高亲和力甲烷氧化活性的发生与恢复
- 与土壤HAMO活性相关的活跃的甲烷氧化菌
- 图2 水稻土中具有和不具有高亲和力甲烷氧化活性的甲烷氧化菌的转录谱
- 已知的甲烷氧化菌的生长和HAMO活性的诱导
- 图3 甲烷氧化菌的丰度与水稻土中高亲和力甲烷氧化活性的出现和恢复有关
- 图4 *pmoA*基因和甲烷氧化菌16S rRNA基因的定量分布
- 土壤HAMO活性的机理及生态意义
- 方法
- 土壤样品的原位描述
- 甲烷冲喂的微宇宙体系构建
- 核酸提取
- 水稻土的宏转录组分析
- 水稻土中活性甲烷氧化菌的稳定同位素示踪
- *pmoA*基因的焦磷酸测序
- *pmoA2*基因的克隆文库构建
- *pmoA*基因的定量
- 16S rRNA基因及其转录本的MiSeq测序
- 杂交技术
- 数据提交
- Reference
- 通讯作者简介
- 猜你喜欢
- 写在后面

传统甲烷氧化菌主导稻田土壤中大气甲烷的氧化
Conventional methanotrophs are responsible for atmospheric methane oxidation in paddy soils
Impact Factor:11.878
https://doi.org/10.1038/ncomms11728
发表日期:2016-06-01
第一作者:蔡元锋1
通讯作者:贾仲君(jia@issas.ac.cn)1
合作作者:郑燕, Paul L.E. Bodelier,Ralf Conrad
主要单位:
1中国科学院南京土壤研究所土壤与可持续农业国家重点实验室(State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing, Jiangsu Province 210008, China.)
写在前面
分享标题:Nature子刊:南土所贾仲君组-揭示水稻土大气甲烷氧化的微生物过程机制
关键字:甲烷氧化菌,CH4,干湿交替,MTWAS,PHB,pmoA基因,稳定同位素示踪
点评:中国科学院南京土壤研究所贾仲君课题组解析了湿地土壤氧化大气甲烷的机制,发现传统甲烷氧化菌,而不是未知的难培养微生物在大气甲烷氧化过程中发挥了主要作用。针对干湿交替的水稻土,课题组提出了全转录组水平的微生物群落关联技术理念MTWAS(metatranscriptome-wide association study),从数以千亿计的遗传信息中,解析了水稻土甲烷氧化过程的微生物代谢调控网络。研究结果表明:高浓度甲烷刺激是水稻土氧化大气低浓度甲烷的前提条件,MTWAS分析则发现传统甲烷氧化菌利用高浓度甲烷过程中,合成了能量物质PHB,极有可能利用PHB氧化大气甲烷,结合高通量功能基因检测和稳定性同位素示踪DNA技术,巧妙排除了难培养大气甲烷氧化菌的贡献。这一结果得到了审稿专家的高度评价,认为是温室气体排放领域的突破性成果、令人惊奇的发现,为全球甲烷和碳循环、气候变化的计算模拟提供了新视角。
摘要
土壤是强温室气体甲烷的生物源,而在大气中只有大约1.84 ppmv(part per million by volume)极低的浓度。
长期以来,人们一直认为这种“高亲和力”甲烷氧化(HAMO)是由尚未被培养的消耗甲烷的细菌造成的。在这里,我们展示了源于稻田土壤中传统甲烷氧化菌的新兴HAMO活性。在高浓度甲烷的低亲和力氧化过程中,很快诱导出HAMO活性。此活性在2周内逐渐消失,但可以通过用高浓度的甲烷来冲喂土壤而重复获得。HAMO活性的诱导是在甲烷氧化菌群快速生长后才发生的,转录组研究表明同时发生的高亲和力和低亲和力甲烷氧化是由已知的甲烷氧化菌催化的,而不是由提出的新型大气甲烷氧化菌催化的。这些结果为定期排水的生态系统吸收大气甲烷提供了证据,这些生态系统通常被认为是大气甲烷的来源。
背景
甲烷(CH4)是仅次于CO2的第二重要的大气温室气体,被认为解释全球变暖的17%。需氧旱地土壤中的微生物会消耗大气中的甲烷,而这些土壤主要暴露于接近或低于大气中甲烷浓度1.84 ppmv的极低水平。土壤甲烷消耗动力学的理论计算表明,大气中的甲烷不能为任何已知的产甲烷生物的存活提供足够的能量支持。现有证据表明,拥有高亲和力甲烷氧化酶(HAMO)的微生物属于尚未被培养的甲烷氧化世谱系。这些谱系由山地土壤群alpha (USCa)和山地土壤群gamma (USCg)组成,这两种土壤类群在好氧山地土壤(如森林和草原地区的土壤)中经常被发现,而在湿地土壤中则没有。
湿地生态系统通常含有高浓度的甲烷,甲烷是有机物厌氧降解的最终产物。在这种情况下,众所周知的低亲和力甲烷氧化菌,包括I型(Gammaproteobacteria)和II型(Alphaproteobacteria)亚群,在好氧厌氧界面催化甲烷氧化。这些界面包括含氧表层土壤和湿地植物释氧根周围的区域。据估计,在厌氧区域产生的内源高浓度甲烷有80%在逸入大气之前被甲烷氧化菌消耗掉。有趣的是,在干旱季节,实地测量表明由于大气中的甲烷以1.84 ppmv的量直接消耗,HAMO活性在一个沼泽区发生,而沼泽土壤在涝渍条件下是甲烷的净来源。在许多其他类型的湿地中,也发现了甲烷源的这种变化,这些湿地水位波动较大,如泥炭地、潮汐淡水湿地和酸性湿地。然而,在这些周期性排干的湿地中,大气甲烷氧化的机制仍不清楚。
稻田可以作为定期排水湿地生态系统的模式系统,因为在亚洲,农业轮作的做法通常用于涝稻种植和旱地小麦种植。此外,稻田的季中排水会由于含水量的变化而导致土壤甲烷浓度的显著波动。因此,我们使用水稻土作为模型系统,通过暴露在不同甲烷混合比例下,模拟周期性排水湿地土壤中甲烷波动的有效性。结果表明,仅在土壤对10,000 ppmv的甲烷氧化后,HAMO的1.84 ppmv甲烷吸收活性才出现,并在2周后停止,但在重新将土壤暴露在10,000 ppmv甲烷中后可以重新获得。尽管甲烷营养型种群的生长逐渐受到限制,但每天向土壤中添加10,000 ppmv甲烷(10天),可以提高HAMO活性的耐受性。整个转录组的关联研究(MTWAS)显示,传统甲烷氧化菌的关键酶具有很强的转录活性,表明多种胞内聚合物可以作为支持土壤HAMO活性的还原剂。我们的研究结果表明,在定期排水的湿地,传统的甲烷氧化菌是造成大气甲烷氧化的原因,而湿地通常被认为是大气甲烷的来源之一。
结果与讨论
土壤中HAMO活性的发生与恢复
Emergence and resilience of soil HAMO activity
我们研究了好氧条件下,水稻土在5种不同混合比例甲烷条件下的甲烷氧化动力学,其包括环境条件下的2 ppmv和高浓度下的100、500、1000和10000 ppmv。在18天的培养期中,暴露于环境空气和100 ppmv甲烷时,土壤中不会发生甲烷氧化(图 1a)。然而,在经过10,000 ppmv甲烷驯化的微宇宙中,甲烷浓度在3天内迅速下降到2 ppmv,并进一步下降到0.6 ppmv(图. 1a),表明土壤中HAMO活性的出现。有趣的是,在500和1000 ppmv甲烷驯化的微宇宙中,虽然顶空甲烷浓度分别下降到10和4 ppmv,但HAMO活性没有被诱导出来。这些结果表明,可能需要临界水平的甲烷转化来诱导稻田土壤中的HAMO活性。
图1 水稻土高亲和力甲烷氧化活性的发生与恢复
Emergence and resilience of high-affinity methane oxidation activity in paddy soil
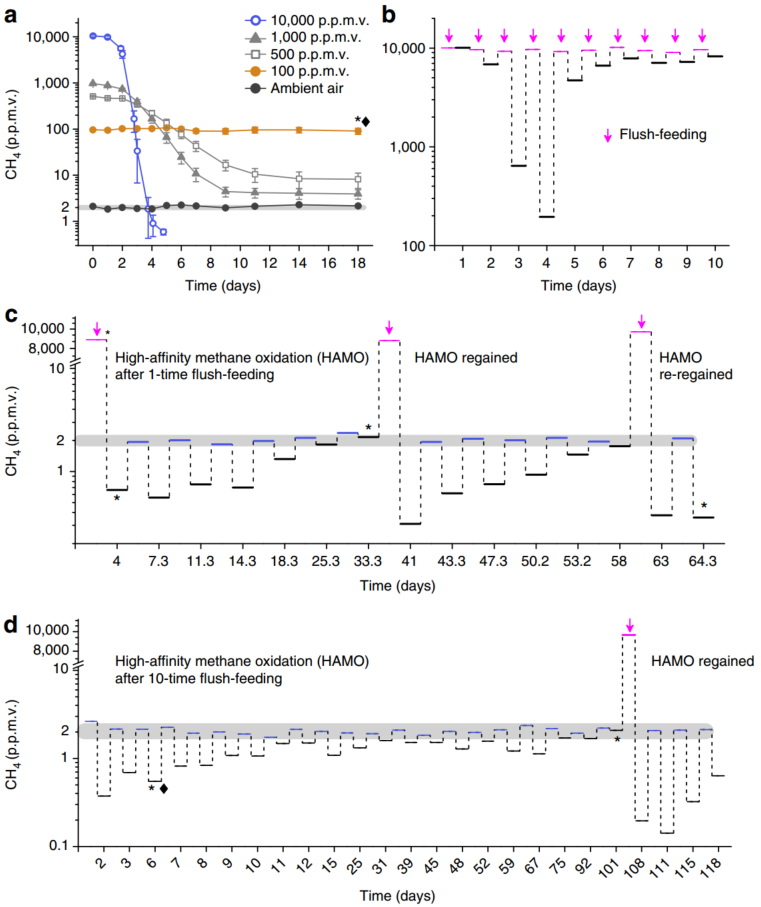
(a)不同初始混合比例下甲烷的消耗动态。高亲和力甲烷氧化(HAMO)活性仅发生在经10,000 ppmv处理的土壤中。误差棒表示从三个微宇宙中测量的上下标准误差。
(b) 在10,000 ppmv甲烷10次冲喂微生物条件下甲烷消耗的变化。顶部空间每天都要更换甲烷,以保持10000 ppmv的甲烷。红色的线条:每次置换后的初始甲烷混合比例; 黑色的线条:培养1天后的甲烷混合比例;
(c,d) HAMO活性在1次(c)和10次(d)冲喂后逐渐丧失和恢复。浅灰色区域代表实验期间大气中甲烷混合比例的范围,黑色线条表示经过数天培养后,土壤微生物中残留的甲烷浓度。a-d图中的黑钻石图标显示了分别用于宏转录组分析的具有和不具有HAMO活性的土壤样品。黑色的星号图标表示用于对pmoA2基因进行定量PCR分析的土壤样本,该pmoA2基因编码了一种对甲烷具有高亲和力新的颗粒状甲烷单加氧酶,并可用于16S rRNA基因和转录本的高通量Illumina测序。b-d中显示的所有数据都是来自一式三份微宇宙的平均值。
为了验证出现HAMO活性的现象并评估其对甲烷转化的依赖性,我们让土壤再次暴露于10,000 ppmv甲烷(1次冲喂 1-TF)中,或者让土壤与10,000 ppmv甲烷共孵育10天(10次冲喂 10-TF),方法是每天更新一次顶空气体(图. 1b)。在10天的孵育期内,经过3-4天的孵育,土壤甲烷消耗量明显增加。然而,随后补充10,000 ppmv甲烷导致甲烷氧化率稳步下降,这表明,由于源于甲烷碳的过度供应,可能会导致营养匮乏(图. 1b)。然后我们用环境空气对这些1-TF和10-TF土壤组进行更新,以确定它们的HAMO活性。值得注意的是,10-TF土壤的HAMO活性并不比1-TF土壤高,尽管前者的甲烷累积氧化量是后者的三倍(图. 1b)。经过三次顶空更新后,1-TF和10-TF土壤中甲烷的消耗动态确实非常相似,表明两种土壤HAMO活性可能受到相似的功能导向和/或受大气甲烷向土壤扩散的限制。经对水稻田中HAMO活性的测定,大气甲烷消耗速率为每年4.46 kg CH 4ha–1 。
但1次冲喂土壤的HAMO活性随时间呈下降趋势,2周后消失(图. 1c)。有趣的是,这种HAMO活性可以在升高的甲烷(10,000 ppmv)孵育下再次被诱导, 尽管随后像以前一样丢失HAMO的活性(图. 1c)。这种活性的重新激活强烈表明,HAMO活性的恢复力和持久性取决于高浓度甲烷的周期性供应和随后的消耗。在10-TF土壤中也观察到了类似的出现和恢复的情景。HAMO活性下降速度与环境空气下的1-TF土壤一样快,但10天后,仍然能检测到微弱的HAMO活性,且能持续>两个月(图. 1d)。值得注意的是,大多数可培养的甲烷氧化菌只有在消耗高浓度的甲烷后才能氧化大气中的甲烷。当以与本研究中暴露的水稻土相似的方式暴露于1.84 ppmv的大气甲烷中时,这种能力通常会在几天内丧失。因此,这表明已知的甲烷氧化菌在周期性排水生态系统的大气甲烷吸收中发挥着重要作用,在这种生态系统中,高甲烷量的冲喂通常伴随着土壤地下水位的波动。
与土壤HAMO活性相关的活跃的甲烷氧化菌
Active methanotrophs associated with soil HAMO activity
我们通过重建整个微生物群落的宏转录组设计了一种MTWAS策略,对HAMO和非HAMO土壤中的目标微生物进行比较分析。在10-TF土壤中,表达的基因和负责HAMO活性的甲烷氧化菌在6天内表现出很强的大气甲烷吸收能力,而在没有HAMO土壤中在18天的孵育期内没有消耗100 ppmv甲烷 (图. 1)。我们使用MG-RAST(Metagenomic Rapid Annotation using Subsystem Technology)服务器对2670万个高质量信使RNA转录本进行了系统分类和功能分类。随后,在水稻土壤甲烷氧化菌基因转录本检测的基础上,重建了甲烷氧化和碳同化的核心代谢途径(图. 2a)。HAMO土壤中存在着编码两种典型甲醛吸收途径的酶基因转录本,I型甲烷氧化菌的一磷酸核酮糖(RuMP)途径,II型甲烷氧化菌的丝氨酸途径。对于甲醛同化的 embdon - meyerhof - parnas(即糖酵解)途径和并行的Enthner–Doudoroff途径在I型甲烷氧化菌中被检测出来。虽然只发现了少数编码6-磷酸葡萄糖酸脱氢酶的gnd基因的转录本,但也发现了甲醛氧化和RuMP再生的异化RuMP循环。我们还观察了与四氢叶酸(H4F)途径相关的基因表达,该途径将C1单位转移到丝氨酸循环中,用于II型甲烷氧化菌的碳同化。我们检测了涉及甲烷经甲醇、甲醛和甲酸氧化成二氧化碳的所有酶反应步骤的基因转录本。如相关与甲醇脱氢酶(由mxaF基因编码)和甲酸脱氢酶(由fdh基因簇编码)的基因转录的高的表达表明,还原当量可由甲醇和甲酸盐的氧化反应持续再生,并供给甲烷单加氧酶(MMO)。MMO可能是推动大气甲烷氧化的关键酶。与预期一样,甲醛在水稻土中通过四氢甲胆碱依赖途径被氧化成甲酸,这与之前对大多数甲烷氧化菌的批量培养研究一致。因此,这些发现表明已知的甲烷氧化菌在田间条件下可能发挥重要作用。
图2 水稻土中具有和不具有高亲和力甲烷氧化活性的甲烷氧化菌的转录谱
Transcript profiles of methanotrophs in paddy soil with and without high-affinity methane oxidation activity
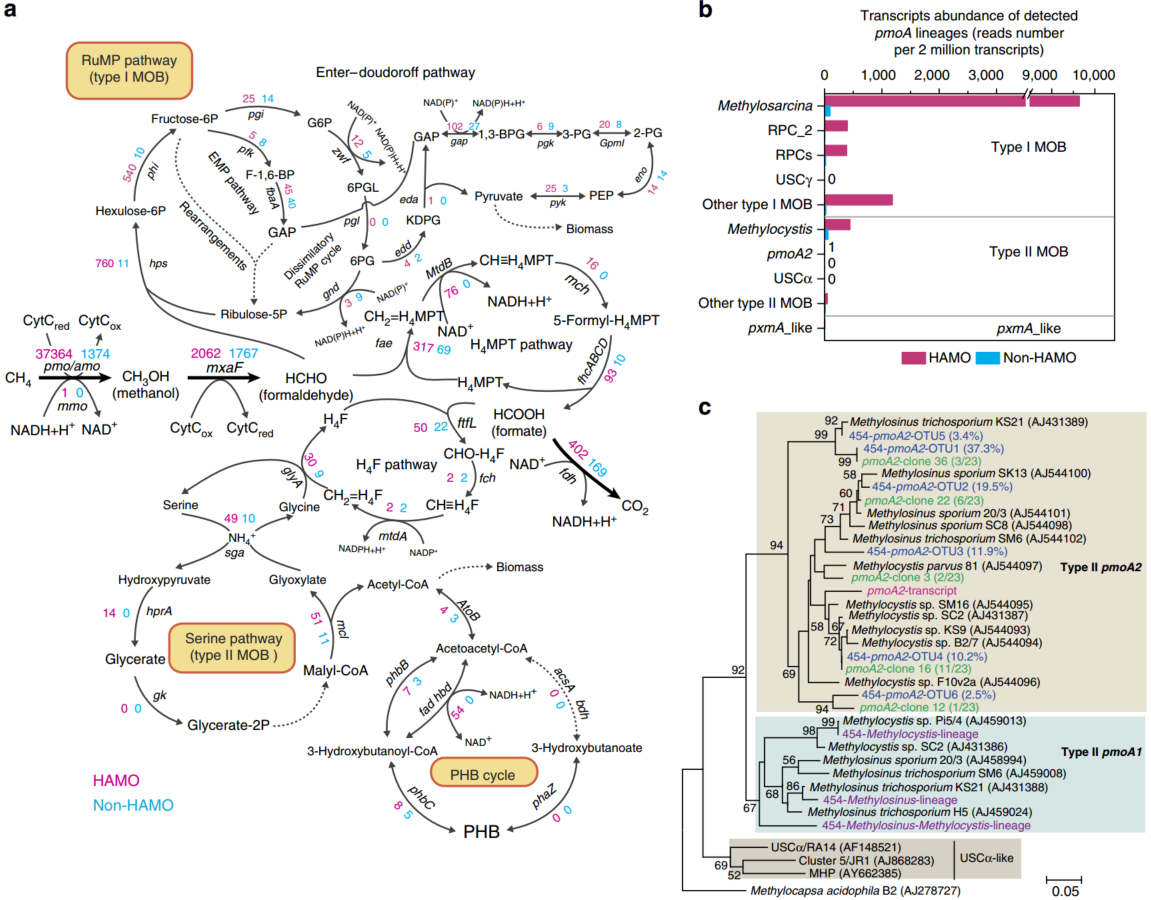
(a)在高亲和甲烷氧化(HAMO)和非HAMO的水稻土中检测到的甲烷氧化菌的甲烷氧化和碳同化途径。基因转录丰度被归一化为每200万个注释转录的读长数。红色和蓝色的数字分别表示在HAMO和非HAMO土壤转录组中检测到的转录丰富度。这些基因编码的酶如下:acsA, 乙酰乙酰基CoA合成酶;atoB,乙酰辅酶a乙酰转移酶;bdh, 3-羟丁酸脱氢酶;eda, 2-脱氢-3-脱氧磷酸葡萄糖酸酯醛缩酶;edd, 磷酸葡萄糖酸脱水酶;eno,烯醇酶; fad, 3-羟烷基-CoA脱氢酶;fae,甲醛活化酶;fbaA,果糖二磷酸酯醛缩酶fch,亚甲基四氢叶酸环水解酶;fdh,甲酸脱氢酶;fhcABCD,甲酰甲基呋喃脱氢酶;ftfL,甲酸四氢叶酸连接酶;gap,3-磷酸甘油醛脱氢酶;gk,D-甘油2-激酶;glyA,丝氨酸羟甲基转移酶; gnd,6-磷酸葡萄糖酸脱氢酶;gpmI,不依赖于2,3-双磷酸甘油酸的磷酸甘油酸突变酶; hbd,3-羟基丁酰基-CoA脱氢酶;hprA,羟基丙酮酸还原酶;hps,3-己糖-6-磷酸合酶; mch,亚甲基四氢甲蝶呤环水解酶; mcl,Malyl-CoA裂解酶;mmo,甲烷单加氧酶(可溶);mtdA,亚甲基四氢叶酸脱氢酶; mtdB,亚甲基四氢甲蝶呤脱氢酶;mxaF,甲醇脱氢酶; pfk,6-磷酸果糖激酶; pgi,6-磷酸葡萄糖异构酶;pgl,6-磷酸葡萄糖酸内酯酶; pgk,磷酸甘油酸酯 激酶;phaZ,聚(3-羟基丁酸酯)解聚酶;phbB,乙酰乙酰辅酶A还原酶;phbC,聚-β-羟基丁酸酯聚合酶; phi,6-磷酸-3-
己糖异构酶;pmo,甲烷单加氧酶(微粒);pyk,丙酮酸激酶;sga,丝氨酸-乙醛酸转氨酶; zwf,6-磷酸葡萄糖脱氢酶。虚线箭头代表多个酶促反应。1,3-BPG,1,3-双磷酸甘油酸酯; EMP途径,Embden–Meyerhof–Parnas途径;F-1,6-BP,果糖1,6-二磷酸; GAP,3-磷酸甘油醛;G6P,6-磷酸葡萄糖; KDPG,2-酮-3-脱氧-6-磷酸葡萄糖酸酯;
MOB,甲烷氧化细菌;PEP,磷酸烯醇丙酮酸; 2-PG,2-磷酸甘油酸酯;3-PG,3-磷酸甘油酸酯; 6PG,6-磷酸葡萄糖酸酯; 6PGL,
6-磷酸-葡萄糖酸-1,5-内酯;PHB,聚-β-羟基丁酸酯; H4F,四氢叶酸; H4MPT,四氢甲蝶呤。(b)HAMO和非HAMO的宏转录组中pmoA谱系的转录丰富度。每个pmoA谱系的转录丰度被标准化为每200万个注释转录的读长数。
(c) 检测到的pmoA2转录本的系统发育(红色)。该树是使用MEGA 4.0基于125个衍生氨基酸序列构建的。pmoA2 OTUs的代表来自pmoA基因扩增子焦磷酸测序(蓝色)和pmoA2基因克隆文库测序(绿色),并且三种pmoA1序列的代表(紫色)也被显示。自展支持率 > 50%(1,000次重复)的在节点上显示。比例条表示5%的置信区间。
与甲烷营养细胞中常见的碳储存聚合物聚-β-羟丁酸(PHB)代谢相关的基因转录本也得到了鉴定(图. 2a)。当细胞受到饥饿、营养缺乏或缺乏还原剂等环境胁迫时,对于甲烷氧化菌来说,PHB代表了一种内源性的还原力来源(例如还原型烟酰胺腺嘌呤二核苷酸NADH)。PHB的积累常发生在碳过剩与氮限制条件下。10-TF土壤中的碳过量可以通过在10天冲喂后期观察到的甲烷消耗的稳步下降得到明确的证明(图. 1b)。固氮基因(固氮酶,由基因簇nifDKH编码)和将细胞外的氨、亚硝酸盐和硝酸盐输送到甲烷氧化菌细胞的基因较高的表达也证明了氮的限制。事实上,我们之前的研究结果表明,在本研究试验的土壤中,当氮约束解除时,甲烷营养化种群和甲烷氧化速率都受到尿素参与的显著刺激。总的来说,高甲烷消耗过程中的诱导应力可能会对甲烷氧化菌细胞的生物量积累产生深远的影响,导致HAMO土壤中富碳化合物的积累(图. 2a)。同时,需要注意的是,PHB循环相关酶的表达可能暗示活性碳的同化。环境应力和PHB积累之间的直接联系还有待实验证实。
在所有甲烷营养基因和基因簇中,编码颗粒状甲烷单氧合酶(pMMO)的pmoCAB是关于HAMO和非HAMO土壤中标准化丰度最丰富的基因簇(图. 2a)。对所有推测的pmoA转录本进行严格的分类显示,在HAMO土壤中,最活跃的甲烷氧化菌谱系为Methylosarcina(79.7%)、稻田集群2(3.2%)和I型甲烷氧化菌的稻田集群(3.2%)和II型甲烷氧化菌的Methylocystis(3.6%)(图. 2b)。12287个pmoA转录本均与被提出的大气甲烷氧化谱系USCa和USCg无关。通过对背景土、非HAMO土、1-TF和10-TF HAMO土的pmoA扩增子进行焦磷酸测序,证实了USCa和USCg谱系的缺失。甲烷氧化菌标志物的深度测序也没有发现在包括中国、印度尼西亚、意大利和越南在内的整个大陆水稻土壤中存在类似USCα和USCγ的pmoA基因的证据。因此,这些新提出的谱系似乎不太可能对水稻土中HAMO活性的出现有贡献(图. 1)。
共441个pmoA转录本可被分配到II型甲烷氧化菌的Methylocystis。这一类群长期以来被认为是大气中的甲烷氧化者。最近的一项研究表明,在菌株Methylocystis sp. SC2中,HAMO活性是由一种新的pmoCAB2基因簇催化的,该基因簇编码了传统pMMO的同工酶(pMMO2)。在HAMO的 宏转录组中也检测到了类似pmoa2的转录本,但在非HAMO土壤中没有检测到(图. 2b)。焦磷酸测序和克隆文库测序进一步揭示了水稻土中系统分化的pmoA2基因(图. 2c),表明具有pMMO2的II型甲烷氧化菌的多样性比以前估计的要大。使用各自的组特异性16SrRNA探针,通过催化报告沉积荧光原位杂交(CARD-FISH)进一步进行细胞可视化。这一分析揭示了典型的I型和II型甲烷氧化菌的独特形态。这一结果提示了它们在水稻土中HAMO活性的发生和恢复中所起的作用。
已知的甲烷氧化菌的生长和HAMO活性的诱导
Growth of known methanotrophs and HAMO activity induction
在HAMO活性的诱导和丢失过程中,通过实时定量PCR (real-time quantitative PCR, qPCR)对pMMO2-拥有型甲烷氧化菌的pmoA2基因进行定量,并通过Illumina MiSeq对所有其他甲烷氧化菌的16S rRNA基因及其一式三份的微宇宙的转录本进行测序。与失去HAMO活性的土壤相比,HAMO土壤中pmoA2基因的拷贝数普遍较高,I型和II型甲烷氧化菌的相对丰度也较高(图. 3)。我们进一步发现,在DNA和RNA水平上,在HAMO土壤中,I型甲烷氧化菌受到的刺激远远大于II型甲烷氧化菌。例如,与时间零点相比,I型甲烷营养体的生物量增加了15.58倍,而II型甲烷营养体在10-TF HAMO土壤中的生物量仅增加了3.95倍(图. 3b,d)。在使用13CH4孵育的10-TF土壤中通过甲烷-C同化的甲烷氧化菌基于DNA的稳定同位素探针(DNA-SIP)进一步证实了该观察结果。
图3 甲烷氧化菌的丰度与水稻土中高亲和力甲烷氧化活性的出现和恢复有关
Abundances of methanotrophs associated with the emergence and resilience of high-affinity methane oxidation activity of paddy soil
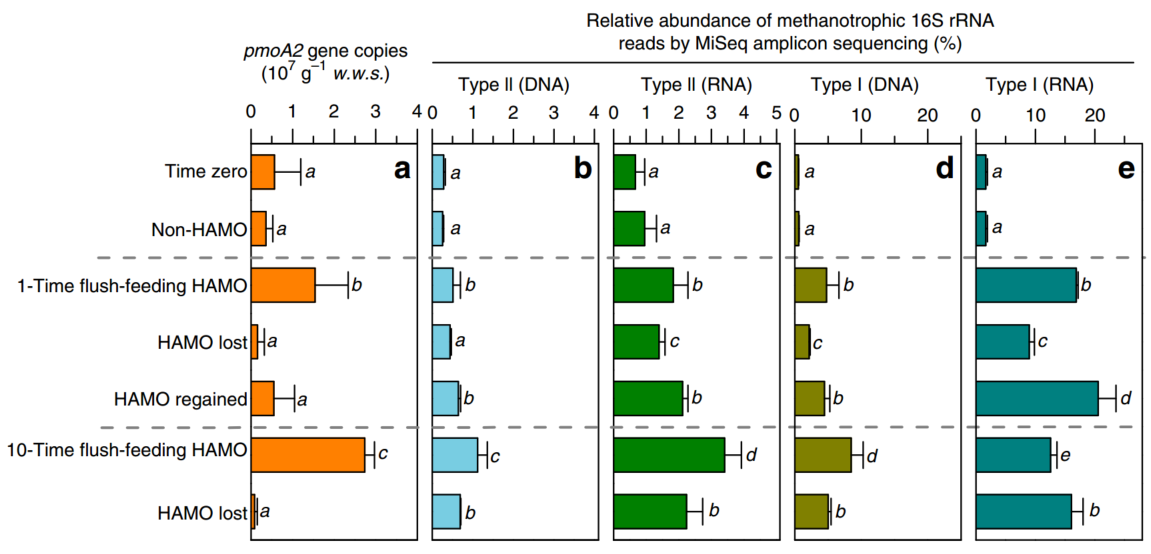
(a)qPCR检测每克湿重土壤(w.w.s.)中pmoA2基因拷贝数(拥有pmoA2基因的甲烷氧化菌)的变化。(b,c) 基于16S rRNA基因(b)和16S rRNA转录本(c)扩增子的Illumina MiSeq测序分析的II型甲烷氧化菌相对丰度的变化。 (d,e) 基于16S rRNA基因(d)和16S rRNA转录本(e)扩增子的Illumina MiSeq测序的I型甲烷氧化菌相对丰度变化。误差棒表示来自一式三份微宇宙测量值的正负误差。柱子上上不同的字母表示有显著差异(方差分析, P<0.05)。
DNA-SIP依赖于生长在13C基质上的活性微生物的细胞分裂。pmoA基因靶向qPCR分析显示,大部分I型甲烷氧化菌是13C标记的,因为它们的基因组DNA是在分离的DNA梯度的重馏分中检测到的(图. 4a)。相比之下,在13C-DNA片段中只检测到一小部分II型甲烷氧化菌和拥有pmoA2基因的甲烷氧化菌(图. 4b,c)。此外,通过对来自每个DNA片段的~130万个高质量的16S rRNA基因读长的分析,我们还发现,在13C标记的DNA中,I型(56.7%)的甲烷氧化菌比例明显高于II型(9.69%)的比例(图. 4d,e)。尽管还不确定生物质碳的积累是源于甲烷基质还是微生物甲烷氧化产生的二氧化碳,这些结果为甲烷氧化菌群落的细胞分裂和繁殖提供了有力的证据。已有研究表明,I型甲烷氧化菌中有5-15%的生物量碳来源于CO2, 对于某些II型甲烷氧化菌,这一数字增加到50%,对于其他的,这一数字增加到62%。此外,如图1b所示,在10000 ppmv时大于90%的13CH4在第3天和第4天被迅速消耗。由此产生的13CO2的量足以支持土壤HAMO活性出现期间的甲烷氧化菌生物量的积累。因此,13C的I型甲烷氧化菌与II型甲烷氧化菌相比,其生长速度更快,这可能在很大程度上代表了之前在该土壤中测试的生命策略的差异,而不是HAMO土壤中碳的基质供应。有趣的是,当1-TF HAMO的土壤在环境空气中培养~2周直至失去活性时,II型甲烷氧化菌的比例仅略有下降(13.1%),而I型甲烷营养体下降了55%(图. 3b,d)。这些结果进一步支持了II型甲烷氧化菌比I型甲烷氧化菌更能耐受环境胁迫(如基质饥饿)的假设,由此推测,II型甲烷营养体可能在HAMO水稻土中发挥重要作用。
图4 pmoA基因和甲烷氧化菌16S rRNA基因的定量分布
Quantitative distributions of the pmoA gene and methanotrophic 16S rRNA gene
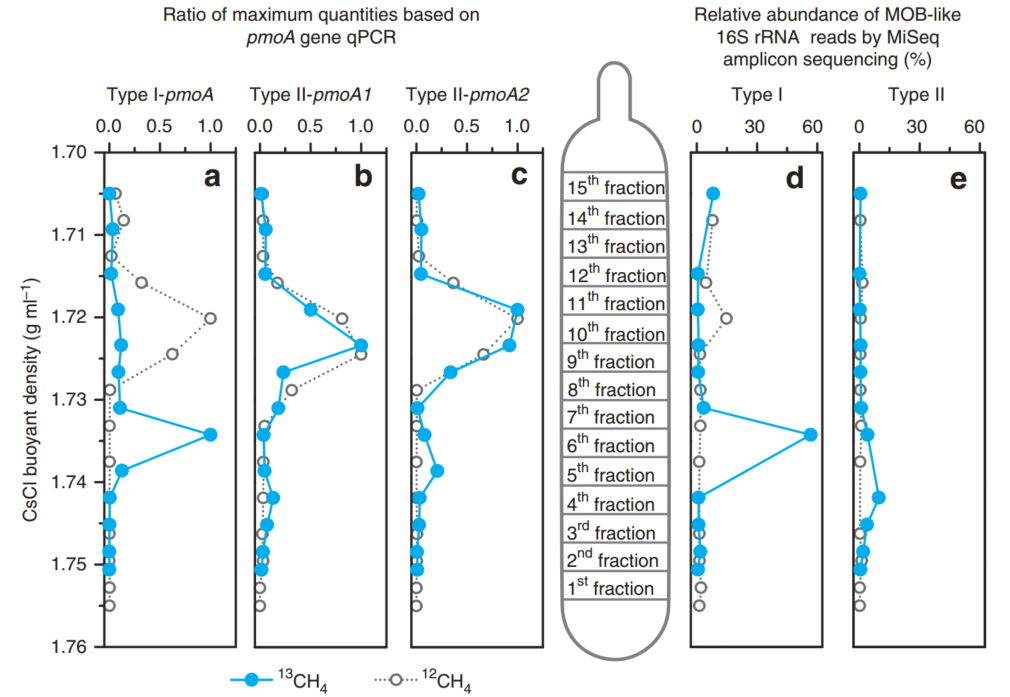
用13CH4和12CH4培养10天HAMO土壤的DNA片段的整个浮密度梯度上都有基因读长。(a–c) pmoA基因在传统I型甲烷氧化菌(a)、传统II型甲烷氧化菌(b)和拥有pmoA2基因的II型甲烷氧化菌中的数量分布(c)。标准化的数据表示为每个片段的基因拷贝数与每个处理的最大数量之比。 (d,e) 基于总16S rRNA基因Illumina MiSeq测序的I(d)型和II (e)型甲烷氧化菌的百分比分布。
已有研究提出,土壤中存在一个诱发HAMO活性的甲烷消耗阈值,本研究可能需要~6.67umol g-1的土壤。土壤微宇宙中表观甲烷氧化速率与顶空甲烷浓度之间存在明显的钟形曲线。我们推测,甲烷氧化菌细胞的快速繁殖可能是在土壤暴露于高浓度甲烷时立即发生的,从而导致土壤甲烷氧化的表观速率越来越高。但是,当每克土壤消耗6.67umol甲烷后,表观氧化速率达到最高值时,最适生长可能会使养分资源迅速减少。在日益增加的压力条件下,甲烷氧化速率开始下降,而细胞生长可能停止,导致富碳化合物在甲烷氧化菌细胞中富集,从而诱导HAMO活性。因此,如果累积消耗量达到每g土壤中6.67 umol甲烷的阈值,那么在浓度为500或1000 ppmv时持续供应甲烷可能会诱发HAMO活性,这似乎是合理的。然而,不同类型土壤的阈值可能有很大差异,其潜在机制仍不清楚。
土壤HAMO活性的机理及生态意义
Mechanism and ecological significance of soil HAMO activity
本研究结果清楚地表明,HAMO活性依赖于高浓度甲烷的频繁供应,这为在自然环境下假设的冲喂机制提供了第一个实验证据。这一理论概念解释了周期性排水湿地中大气甲烷氧化的现象。我们认为,由于地下水位下降,淹水期厌氧聚合生态位内产生的高浓度甲烷会缓慢向上扩散,促进表层土壤好氧/厌氧界面的甲烷氧化菌快速生长。这种低亲和力的消耗进一步促发了干旱季节(当排干的需氧土壤暴露于周围空气中时)甲烷氧化菌吸收大气甲烷以实现高亲和力的HAMO活性。本研究评估稻田土壤的低、高亲和性,为定期排水湿地大气甲烷氧化提供了机制依据。根据本研究观察到的HAMO活性,考虑到全世界有4.1亿公顷的稻田定期进行排水管理,大气甲烷的汇强度估计每年高达40万吨。
水稻土中的HAMO活性很可能是在低养分、胁迫诱导条件下获得的。在10天的培育期中,每天冲喂10,000 ppmv甲烷一次,在第4天激发了显著的甲烷氧化活性。然而,从第5天开始,这种低亲和氧化反应就大大减慢了,这表明甲烷氧化菌的功能逐渐受到损失。同时,由于土壤中氮的限制和氧的消耗,生态位恶化的过程也可能严重加剧。因此,10-TF土壤中甲烷氧化菌的快速生长所产生的环境胁迫可能比1-TF土壤(图. 1a,b)强得多,导致了还原剂(如 NADH)和PHB的积累量增加。这一结果可以解释在10-TF土壤和1-TF土壤中,>2个月的HAMO活性更强(图. 1c,d)。这些内源储备物质长期以来被认为是MMO酶支持培养的甲烷氧化菌HAMO活性的还原力。事实上,HAMO活性的丧失常常被解释为还原力的耗尽。
MTWAS结果揭示了II型甲烷氧化菌中与PHB代谢相关基因的转录活性,表明它们对HAMO活性的诱导具有重要作用。越来越多的证据表明,II型甲烷营养生物比其I型甲烷氧化菌更贫营养,这可能赋予了前者储存化合物合成的优势。有趣的是,几乎所有II型甲烷氧化菌的pmoA转录本在系统发育上都与Methylocystis密切相关,而Methylocystis是迄今为止最贫营养的甲烷氧化菌。尽管在如此低的甲烷浓度下没有观察到生长,但是某些Methylocystis菌株甚至可以在大气中氧化甲烷长达3个月。最近的宏分析表明,在周期性排水的生态系统和高地土壤中普遍存在Methylocystis,在其田间土壤中经常检测到大气甲烷氧化。在本研究中,HAMO活性的维持可能部分解释为pmoA2基因的表达,该基因仅存在于HAMO土壤中 (图. 2c)。重要的是,该基因对甲烷饥饿条件下的Methylocystis的生存至关重要。应该指出的是,一些Methylocystis菌株可以使用醋酸盐作为碳源,醋酸盐氧化产生的还原力可以维持HAMO活性。
I型甲烷氧化菌也可能在大气甲烷氧化中起作用。在HAMO的宏转录组中观察到的pmoA转录本数目非常高,高达9638个,表明与Methylosarcina具有高度的序列相似性。几种I型的甲烷氧化菌已经显示出在大气水平下氧化甲烷的短暂能力。然而,I型甲烷氧化菌被广泛认为是竞争者,其pmoA转录本很可能是在HAMO土壤中快速消耗高浓度甲烷的过程中合成的。Methylosarcina物种是否能消耗大气中的甲烷也是未知的。同时,在已知的I型甲烷氧化菌中,PHB合成的基因组证据仍然缺失。然而,不能排除的可能性是,其他储存化合物可能用于支持I型甲烷氧化菌中的HAMO活性。例如,在HAMO转录组中检测到糖原代谢的基因转录本。考虑到所有被测试的I型甲烷氧化菌都储存糖原,它们也应该能够在饥饿条件下通过使用这种储存化合物向pMMO提供还原剂。此外,如纯培养所示,中间代谢物,如甲醇或甲酸盐,也可作为还原剂来维持HAMO活性。因此,在高分类学分辨率下使用选择性抑制剂来区分甲烷氧化菌,可能对量化不同已知甲烷氧化菌对土壤中大气甲烷氧化的相对贡献有很大的前景。
总之,我们的结果表明,只有在水稻土中高浓度的甲烷被消耗后,才能诱导大气中的甲烷才被氧化。然而,除非土壤定期被高浓度的甲烷冲刷,否则诱导活性就会丧失。在耗尽冲喂的甲烷时,甲烷氧化菌可能依赖于内源性存储化合物(例如PHB)作为共同底物来维持细胞活力,同时消耗大气中的甲烷。水稻土中同时发生的高亲和力和低亲和力的甲烷氧化是由已知甲烷氧化微生物菌群催化的,而不是USCa和USCg中假定的大气甲烷氧化菌群。水稻土中HAMO活性的出现和恢复为更好地理解周期性排水生态系统中的甲烷转化提供了机制基础,并且有必要重新评估陆地生态系统中的大气甲烷氧化,以减少全球甲烷预算的不确定性。
方法
土壤样品的原位描述
Site description of the soil samples
土壤样本取自江苏省扬州市中国典型亚热带农业区的一块主要用于水稻和小麦轮作的水稻田。水稻收获后,用土钻收集3个重复的土壤芯(深度为0-15厘米),过2毫米的筛网进行混匀,然后在4摄氏度下保存,直到进一步使用。土壤特征如下:最大持水量,55%;总有机碳,15g kg-1;总氮,1.59 g kg-1;总P, 1.23 g kg-1;pH 7.4。
甲烷冲喂的微宇宙体系构建
Microcosm construction with the flush-feeding of methane
微宇宙体系建造之前,于环境空气条件下,土壤在60%持水量和28 °C的黑暗环境中培养4天。然后,将0.5 g湿重的土壤放入无菌的2毫升含有1ml RNAlater (Ambion)离心管中,作为初始样品(时间记为0时刻),保存在-20℃下,直到核酸提取。如前所述,将5g预培养的水稻土加入到120 ml的血清小瓶中,盖上丁基橡胶塞子。分别以高浓度甲烷进行1-TF和10-TF处理。1-TF微宇宙的初始甲烷浓度为10,000、1,000、500、100和2 ppmv(环境空气)。在10-TF微宇宙中,10,000 ppmv的 CH4每天更新一次,持续10天。用气相色谱-火焰离子化检测器(GC-FID)(Shimadzu GC12-A, Japan)测定了每瓶顶空的甲烷浓度。HAMO活性指的是在微宇宙中顶空甲烷在2 ppmv以下的消耗。顶空气体用环境空气冲洗以监测HAMO活性的持续性。当土壤不能氧化大气中的甲烷时,将其再次暴露于10,000 ppmv的CH4中,测定HAMO活性三次。在用环境空气冲洗顶空气体后,采用与1-TF土相同的方法对10-TF土的HAMO活性进行了监测。当土壤获得或失去HAMO活性时,进行破坏性土壤取样。实验期间还对装有10,000 ppmv CH4的空瓶进行了监测,以确保没有气体泄漏。所有处理至少进行了三次重复。
水稻土的HAMO活性是根据前3或3.5 h可被氧化的大气CH4含量来评估的。120ml瓶中CH4浓度下降速率为0.18584 ppmv h-1(1-TF和10-TF处理的平均值),土壤覆盖面积为~1.256*10 -3 m2,深度为
~0.5厘米。假设在稻田中,可以诱导2 cm表层土壤的HAMO活性,在25℃,1 atm的压力下甲烷密度为0.7163 gl -1。据估计,HAMO土壤的大气甲烷氧化速率可能高达每天每平方米305.22 mg,相当于每年4.46 kg ha-1。
核酸提取
Nucleic acid isolation
如前所述(稍加修改),从土壤样品中提取总核酸含量。简而言之,土壤样品储存在-20℃的RNAlater中,在冰上解冻,重悬,在20000 g的条件下离心聚集1分钟。然后将离心产物与0.5 g玻璃珠混合,用酸性裂解缓冲液进行两轮细胞裂解。采用水饱和酚(pH 4.5)、酚氯仿异戊醇(25:24:1, (vol/vol/vol), pH 4.5) 、氯仿异戊醇(24:1, (vol/vol), pH
5.5) 依次提取上清。使用两倍体积的PEG NaCl沉淀核酸,用400 ml 70%乙醇洗涤,重悬于50 ml无核酸酶的H2O中,其中10 ul稀释10倍,用于基于DNA的扩增。对于RNA分离,其余的提取物用重组DNase I(Takara)处理以消化DNA,然后使用RNeasy Mini Kit(Qiagen, Germany)纯化。采用以纯化RNA为模板的通用16S rRNA基因引物(515F, 50-GTGCCAGC
MGCCGCGG-30; 907R, 50-CCGTCAATTCMTTTRAGTTT-30)进行PCR扩增以排除DNA污染。使用PrimeScript 1链cDNA合成试剂盒(TaKaRa)和随机六聚体将总RNA转化为互补DNA (cDNA),并保存于-20°C用于扩增子测序。
水稻土的宏转录组分析
Metatranscriptomic analysis of the paddy soil
从HAMO(10-TF土壤,6天内表现出强烈的的大气甲烷吸收)和非HAMO(经100 ppmv甲烷修饰的土壤,在18天的潜伏期内没有甲烷氧化)土壤(如图1所示)中分别获得了宏转录组。按照上述方法从一式三份的微宇宙中提取每个土壤样品的总RNA,并将其汇集在一起。然后,使用Ribo-Zero Magnetic Kit试剂盒(Bacteria; Epicentre)将rRNA去除。使用Superscript Double-Stranded cDNA Synthesis kit(Invitrogen)和随机六聚体引物获得cDNA,然后使用Covaris M220将其裂解为~200 bp。文库使用TruSeq DNA Sample Prep Kit构建,测序在Illumina HiSeq2000平台进行。
原始FASTQ数据文件被提交到MG-RAST 3.0进行分类和功能注释。信使RNA转录本的分类学隶属关系是使用M5NR蛋白数据库中的“Best Hit Classification”方法确定的,参数如下:平均e值小于或等于1e-5,平均比对长度≥30 aa和平均%identity ≥60。将隶属于甲烷氧化菌的转录本选择到workbench中,然后使用Subsystem 数据库对这些转录本(平均e值≤1e-10,平均比对长度≥30 aa和平均%identity ≥60)进行功能注释。在重建甲烷和氮的代谢途径之前,将HAMO和非HAMO宏转录组中每个功能类别的转录本丰度标准化为Subsystem数据库注释的每百万总转录本中的读长数。
我们选择了HAMO和非HAMO 宏转录组中所有假定的pmoA基因转录本来推断活性甲烷氧化菌的分类特征。这个过程是这样进行的。(1)如前所述,我们已经完成了一个包含6,628个可培养甲烷氧化菌株和尚未培养甲烷氧化生态型(如USCa和USCg)的pmoA和pmoA相关序列数据库的准备工作。(2)完成了土壤中pmoA基因转录本的筛选。首先利用GenBank nr数据库作为注释源,对MG-RAST中两个宏基因组进行基因功能注释(e值≤1e-5,平均比对长度≥30 aa和平均%identity ≥60);然后,选择所有假定的pmoA序列(功能注释中含有“甲烷单加氧酶”)并以fasta格式下载。(3)根据pmoA数据库(步骤1)对假定的pmoA序列进行BLAST搜索,然后使用前面所述的MEGAN中最低共同祖先法将其分类为已知的pmoA谱系。将HAMO和non-HAMO宏转录组中每个pmoA谱系的转录本丰度标准化为使用GenBank nr数据库注释的每200万个转录本中的读长数。
水稻土中活性甲烷氧化菌的稳定同位素示踪
Stable isotope probing of active methanotrophs in paddy soil
为了监测活跃的甲烷氧化菌在HAMO活性出现和恢复期间的活动情况,分别用10,000 ppmv ,13CH4和12CH4的DNA-SIP孵育作为标记处理和对照处理。在10-TF微宇宙中进行破坏性采样。使用FastDNA spin kit (MP Biomedicals, Cleveland, OH, USA)根据制造商的说明对土壤提取基因组DNA。DNA-SIP分馏按前面描述的方式进行。生成多达14个梯度部分,并使用AR200数字手持折射计(Reichert, Inc., Buffalo, NY, USA)测定每个部分的折射率。分离的DNA经PEG-6000沉淀法回收,于30 ml TE缓冲液中重悬,用于MiSeq测序和real-time qPCR。
pmoA基因的焦磷酸测序
Pyrosequencing of pmoA gene
利用pmoA基因的多重扩增子焦磷酸测序技术对水稻土甲烷氧化群落进行了综合评估。选取time zero、non-HAMO、1-TF HAMO和10-TF HAMO土壤的DNA样本,利用barcode引物A189f/A682r进行PCR扩增。将来自一式三份微宇宙汇聚的扩增子按等摩尔比混合,并在Roche 454 GS FLX Titanium sequencer(Roche)上进行测序。通过Mothur软件(版本1.33.3)使用‘trim.seqs’和‘split groups’命令处理原始序列文件,用于质控和分样。然后,使用在线版本的FunGene Pipeline对这些读长进行处理,使用USEARCH 6.0检查chimera,使用FramBot对frameshifts进行校正。每个样本的高质量pmoA序列被归类为上述已知的pmoA 群或谱系。选取某一特定类群或谱系的pmoA序列,从MEGAN处下载,基于通过距离cutoff值为0.07的FunGene流程将其推断的氨基酸序列聚类成种级操作分类单元(OTU)。提取每个谱系(各谱系优势OTU的代表)的一个代表和pmoA2基因主要OTU的几个代表,并将其用于系统发育分析。通过具有1,000个重复引导的MEGA 4.0利用neighbour-joining方法建立了pmoA的系统发育树。
pmoA2基因的克隆文库构建
Clone library construction of the pmoA2 gene
在10-TF土壤中也构建了pmoA2基因的克隆文库。如前所述,pmoA206f/pmoA703b引物对用于扩增pmoA2基因。将3个重复扩增子汇集到一起,连接到pGEM-T载体(Promega, Fitchburg,
WI, USA) 上,并转化进合格的DH5α感受态细胞中。随机选取阳性克隆进行Sanger测序,获得23条pmoA2序列。通过FunGene流程进行OTU聚类和代表性序列的挑选。
pmoA基因的定量
Quantification of the pmoA gene
如前所述,利用引物对A189F/Mb601R和II223F/II646R,通过qPCR对片段DNA中I型和II型甲烷氧化菌的常规pmoA基因丰度进行定量。利用引物对pmoA206f/pmoA703b测定了不同甲烷处理土壤和片段DNA中pmoA2基因的拷贝数。反应在20 ul混合物中进行,其含10.0ul SYBR Premix Ex Taq (Takara)和每个引物0.5mM及1ulDNA模板。所有qPCR检测(一式三份)均在CFX96 Optical Real-Time Detection System (Bio-Rad, Laboratories Inc., Hercules, CA, USA)上进行。分别用包含有相应pmoA基因片段的质粒进行系列10倍稀释得到相应的标准品。三对引物的扩增效率均在81.5 ~ 94.1%之间,R2值为0.993 ~ 0.998。
16S rRNA基因及其转录本的MiSeq测序
MiSeq sequencing of the 16S rRNA genes and their transcripts
通过Illumina MiSeq对来自三个重复微宇宙的16S rRNA基因和转录本进行测序,评估了不同采样点的微生物群落结构。还对DNA-SIP各组分的微生物群落组成进行了评估。515F / 907R引物对被用于扩增16S rRNA基因的V4-V5区,并将12 bp的特异性样品接头序列连接到515F的5’端。使用TaKaRa
Taq Hot Start Version (TaKaRa, Japan) 以50ul的总体积包含1.5U TaKaRa Taq HS,4 ul dNTP混合物(10 mM),5ul 10* PCR缓冲液,0.5ul牛血清白蛋白(20mg ml -1Takara),每种引物1ul(10 mM)和1ul模板DNA进行PCR。PCR条件为:94℃2 min;30个循环,包括94℃30秒、60℃30秒和72℃45秒;随着最后一步72 °C5分钟的延伸后,所有扩增子均在经Goldview染色的1.2%(W/V)的琼脂糖凝胶上进行验证,并使用MiniBEST DNA片段纯化试剂盒3.0版(TaKaRa)进行纯化,用NanoDropND-1000分光光度计定量,并按等摩尔比混合。
使用TruSeq Nano DNA LT Sample Prep Kit Set A (24个样品)构建文库,并按照制造商的方案使用MiSeq Reagent Kit
v3(600个循环)进行测序。
杂交技术
CARD-FISH
我们使用杂交技术来识别I型和II型甲烷氧化菌,以及前面描述的总细菌。如先前描述的进行一些修改,从1.0g水稻土中一式三份提取细胞。简而言之,首先将土壤悬浮于10 ml 0.2%焦磷酸钠中,并通过涡旋振荡15分钟使其匀浆,然后将土壤匀浆铺在相同体积的饱和蔗糖溶液(1.33g ml-1)上。5500 g离心2分钟后,将含有细菌细胞的上层蔗糖级分倒入新的15ml聚异丁烯离心管中,用1/3体积的0.8%NaCl稀释,并在4°C下于12,000g离心10分钟。然后用4ml 0.8% NaCl重悬细胞聚集物,转移到含有4ml Nycodenz溶液(1.310g ml-1)的10ml灭菌管中。在4°C下以9,500g离心25分钟后,回收Nycodenz和水层之间的细胞层,洗涤,然后在4°C下以12,000g离心10分钟使其沉淀。用0.2ml PBS重悬细胞沉淀,用于杂交分析和4,6-二氨基-2-苯基吲哚染色。简而言之,在25℃下用甲醛(2%的最终浓度)固定45分钟后,将细胞悬浮液过滤到0.22mm的聚碳酸酯膜(Millipore)上。然后,将滤膜用1倍的PBS洗涤,风干,在37℃下用溶菌酶(10mg / ml 1)处理30分钟,并再次用水洗涤。
然后将滤膜与辣根过氧化物酶标记的探针在杂交溶液中于46°C杂交3小时。用探针EUB I-III、Mγ669和Mα450分别对总细菌和I型和II型甲烷氧化菌进行16S靶向rRNA杂交。探针NON338作为阴性对照。然后将滤膜用洗涤缓冲液洗涤(在48°C下30分钟),转移到含有0.0015%H2O2和1mg ml-1 Oregon Green标记的乙酰胺的扩增缓冲液中,在46 C下洗涤30 min,并用1*PBS洗涤 并在25℃,黑暗中用4,6-二mid基-2-苯基吲哚(50ng ml-1)染色10分钟。在滤膜包埋在Citifluor / Vectashield混合物中后,用落射荧光显微镜下观察滤膜并拍照(Nikon Eclipse 80i)。
数据提交
Data availability
所有支持本研究结果的序列数据已存入以下公共数据库。16SrRNA和pmoA基因的原始扩增子序列数据集已根据登录号ERP008876存入European Nucleotide Archive(ENA)。转录组数据已存储在MG-RAST中,其登录号为4553284.3和4553285.3。从克隆文库测序获得的所有pmoA2基因序列均已存入GenBank,登录号为KP218949至KP218971。图(图2-4)的源数据随文章提供在补充信息文件中。作者声明所有支持本研究结果的相关数据均可在文章中获得,并可根据要求提供补充信息。
编译:马腾飞 南京农业大学 莫永亮 中科院南土所
责编:刘永鑫 中科院遗传发育所
Reference
Yuanfeng Cai, Yan Zheng, Paul L.E. Bodelier, Ralf Conrad,Zhongjun Jia. Conventional methanotrophs are responsible for atmospheric methane oxidation in paddy soils.Nature Communications volume 7, Article number: 11728 (2016) doi: https://doi.org/10.1038/ncomms11728
通讯作者简介

贾仲君,中科院南京土壤研究所研究员、博士生导师。1997年毕业于山西大学; 2002年于南京土壤研究所获博士学位。2002-2003研究草地土壤大气甲烷氧化微生物(美国农业部与科罗拉多矿业大学);2003-2006年从事稻田甲烷和氨氧化微生物生理生态学研究(日本名古屋大学);2006-2008年从事土壤氨氧化微生物生理生态学研究(马普陆地微生物研究)。2008年入选中国科学院引进国外杰出人才项目百人计划。从事土壤环境微生物研究。采用稳定同位素原位标记土壤微生物核酸DNA的先进手段,在国际上首次提出证据表明氨氧化细菌,而非氨氧化古菌是农田土壤氨氧化过程的主要驱动者;在国际上首次发现非水生环境(土壤)存在特异的T4噬菌体;稻田生态系统甲烷氧化和氨氧化既可能相互促进,也可能相互抑制。采用荧光实时定量PCR标靶功能基因技术,显著提高了13C-DNA检测灵敏度,发现氨氧化细菌AOB主导德国典型农田土壤氨氧化过程;进一步开发了新一代454高通量测序为基础的DNA-SIP技术,初步定量了氨氧化细菌AOB和氨氧化古菌AOA对农田土壤硝化过程的相对贡献,表明AOB主导我国华北平原典型潮土氨氧化;结合硝化抑制剂技术,清楚揭示了复杂农田土壤中硝化微生物群落无机化能自养生长的生理代谢特征。相关研究成果发表在国际知名刊物如Nature Communications ,Environmental Microbiology, The ISME Journal等杂志。曾获国际全球变化分析研究培训系统总部(START)杰出青年科学家奖,中国科学院院长优秀奖,江苏省科技进步一等奖,江苏省“333高层次人才培养工程”培养对象荣誉称号。
猜你喜欢
- 10000+: 菌群分析
宝宝与猫狗 提DNA发Nature 实验分析谁对结果影响大 Cell微生物专刊 肠道指挥大脑 - 系列教程:微生物组入门 Biostar 微生物组 宏基因组
- 专业技能:生信宝典 学术图表 高分文章 不可或缺的人
- 一文读懂:宏基因组 寄生虫益处 进化树
- 必备技能:提问 搜索 Endnote
- 文献阅读 热心肠 SemanticScholar Geenmedical
- 扩增子分析:图表解读 分析流程 统计绘图
- 16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
- 在线工具:16S预测培养基 生信绘图
- 科研经验:云笔记 云协作 公众号
- 编程模板: Shell R Perl
- 生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习扩增子、宏基因组科研思路和分析实战,关注“宏基因组”


点击阅读原文,跳转最新文章目录阅读
https://mp.weixin.qq.com/s/5jQspEvH5_4Xmart22gjMA
最后
以上就是内向哈密瓜最近收集整理的关于Nature子刊:南土所贾仲君组-稻田甲烷氧化的微生物机制传统甲烷氧化菌主导稻田土壤中大气甲烷的氧化的全部内容,更多相关Nature子刊:南土所贾仲君组-稻田甲烷氧化内容请搜索靠谱客的其他文章。








发表评论 取消回复